Биология
Грибы порядка Erysiphales – облигатные паразиты цветковых растений. В основном поражают двудольные. Из однодольных, растениями-хозяевами мучнисторосяных грибов, являются различные виды злаков. В цикле развития патогенов порядка присутствуют конидиальная стадия (анаморфа) и сумчатая стадия (телеоморфа).
Сумчатая стадия (телеоморфа)
Сумчатая стадия возникает в результате протекания полового процесса. Развитие наблюдается обычно в конце вегетационного периода. На мицелии образуются мужские и женские половые органы. Они устроены просто. Мужские (антеридии) состоят из двух клеток. Женские (аскогоны) – из одной. Содержимое верхней клетки антеридия через специальное отверстие переливается в аскогон. При этом происходит оплодотворение. После этого аскогон разрастается и делится на несколько клеток. Одна из клеток получает два ядра (дикарион). В односумчатых плодовых телах оплодотворенная клетка трансформируется в аску. В многосумчатых – из оплодотворенной клетки вырастают аскогенные гифы, на каждой из них развивается аска. Позднее они обрастают гифами мицелия, образующим два слоя перидия. Наружный, состоящий из плотных, темно окрашенных гиф, представляет оболочку. Внутренний – состоит из быстро лизирующихся гиф и выполняет питательные функции.
При созревании асков плодовые тела растрескиваются, аски разрываются и аскоспоры освобождаются. Они попадают на восприимчивые органы растений, прорастают и вызывают заражение. Созревание плодовых тел у большинства видов наблюдается весной или поздней осенью. С экологической точки зрения плодовые тела мучнисторосяных грибов служат для перенесения грибами неблагоприятных условий во время перезимовки.
Конидиальная стадия (анаморфа)
Конидиальная стадия формируется после прорастания аскоспор и состоит их мицелия и конидиального спороношения. На поверхности или внутри зараженных органов развивается мицелий, на нем конидиеносцы с конидиями.
Морфология
Анаморфа (конидиальная стадия)
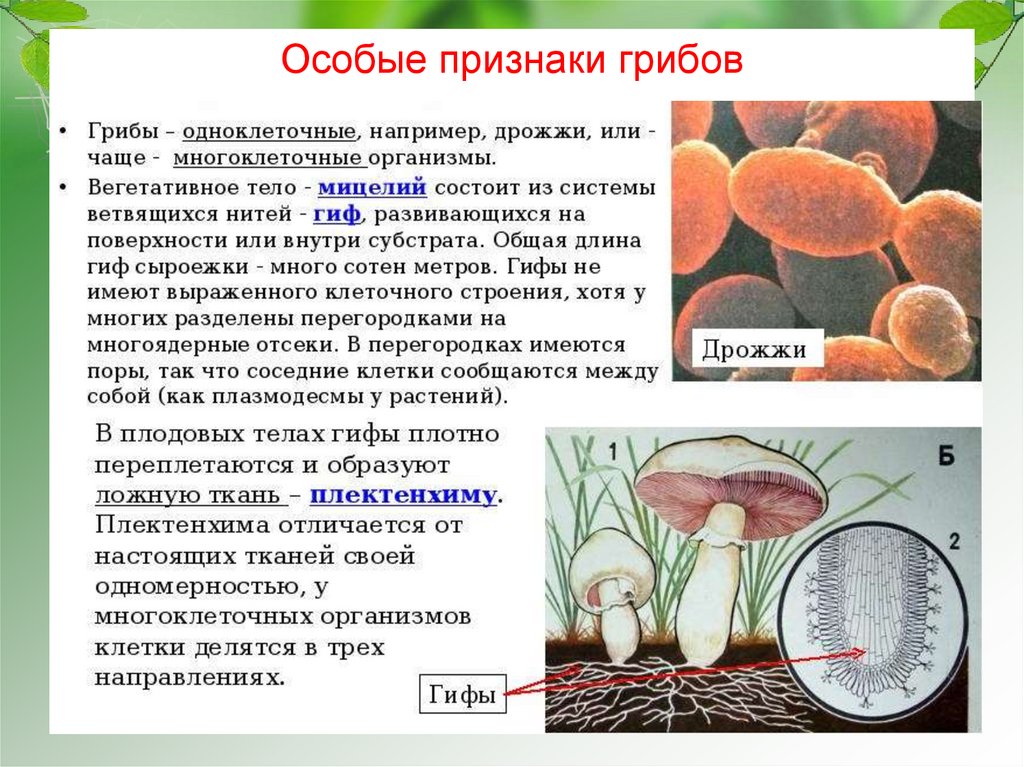
Мицелий – эпифитный, полуэндофитный, эндофитный.
Эпифитный – развивается на поверхности пораженных органов, имеет вид беловатого паутинистого или войлочного налета. Представлен неокрашенными, септированными, тонкостенными гифами, которые состоят из одноядерных клеток.
Полуэндофитный – поверхностные гифы, проникают внутрь листовой пластины через устьица, как у рода Phyllactinia.
Эндофитный – межклеточный мицелий, является вторичным и образуется при переходе вида к конидиальному спороношению. Характерен для представителей рода Leveillula.
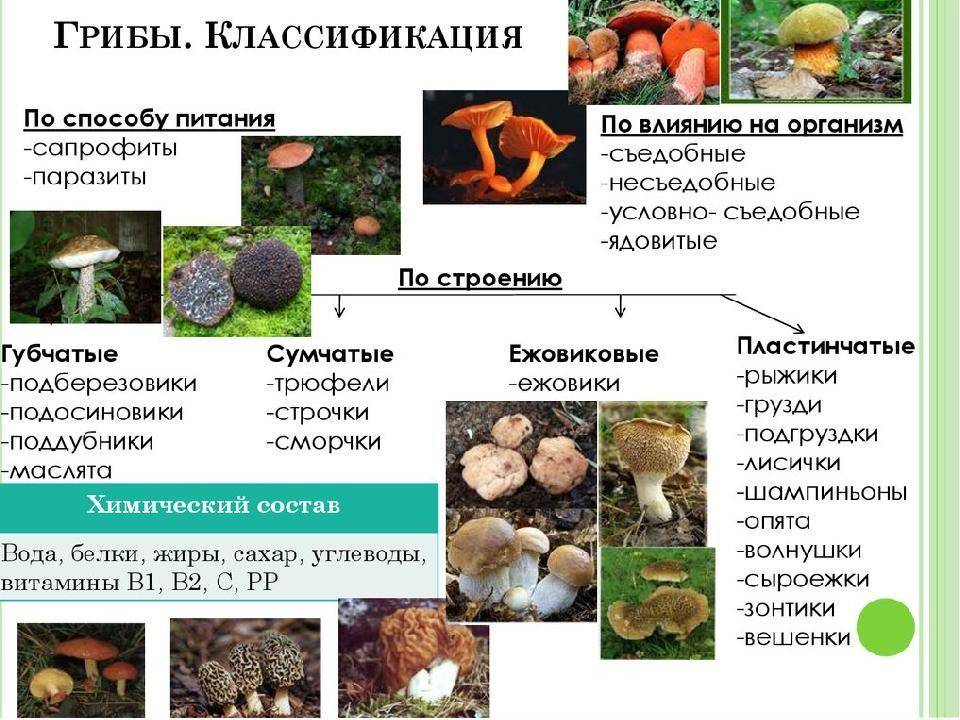



На гифах мицелия формируются инфекционные выросты, проникающие через стенку в клетки эпидермы хозяина и формирующие в ней гаустории.
Гаустории – в основном округлые или мешковидные, у Blumeria graminis – пальчатые. В молодом состоянии гаустории без отростков, в зрелом – образуется различное количество отростков. У всех мучнисторосяных грибов гаустории выполняют две функции: всасывания питательных веществ из клетки эпидермы растения и прикрепления мицелиальной гифы к поверхности тканей. У некоторых видов функцию прикрепления выполняют апрессории (специализированные выросты клеток мицелия).
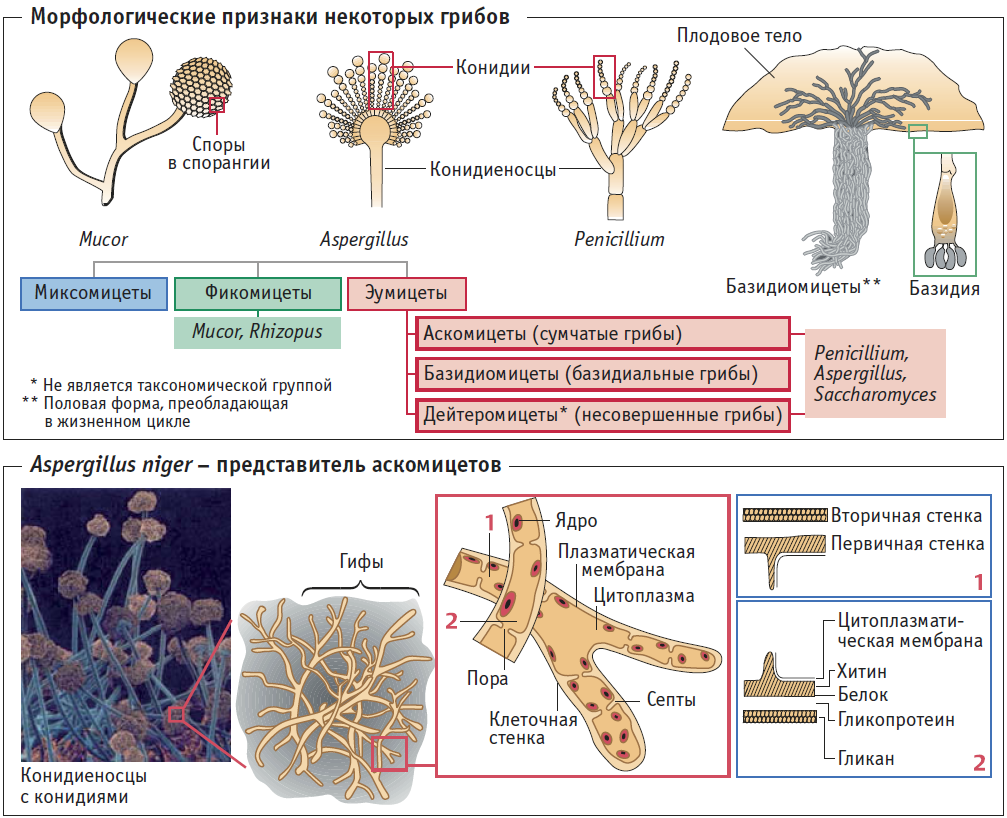
Конидиеносцы – возникают, как отростки или ответвления на гифах мицелия. Постепенно они нарастают и образуют септы.
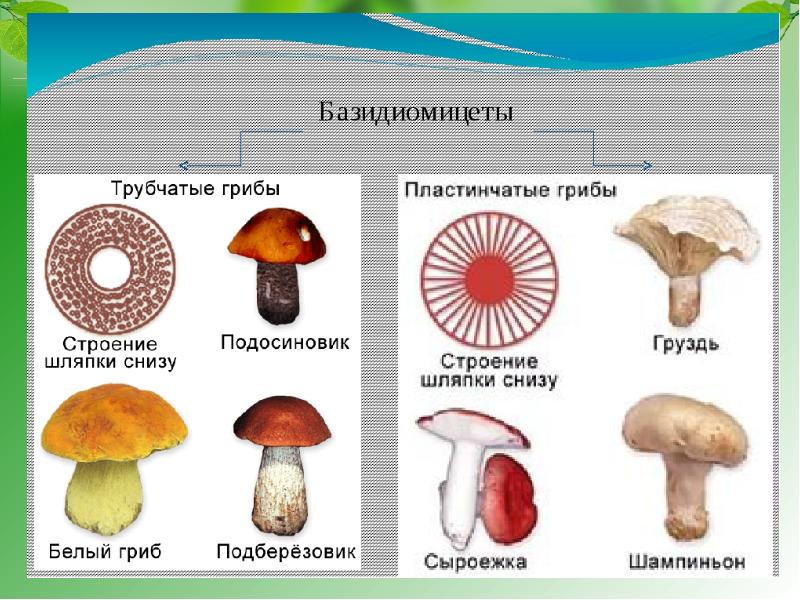
Конидии – по типу образования являются меристем-артроконидиями. Часто образуют цепочки, могут быть одиночными. По форме – цилиндрические, эллипсоидальные, бочковидные, ланцетовидные. Конидии придают пораженным органам растений «мучнистый вид» характерный для заболеваний, вызываемых мучнисторосяными грибами.
Телеоморфа (сумчатая стадия)
Плодовые тела – замкнутые, аски расположены пучком, аскоспоры разбрасывают активно. Плодовые тела видов порядка Erysiphales рассматривают как перитеции или как клейстотеции. В целом их считают переходной формой от клейстотеция к перитецию, иногда их называют клейстокарпии.
На поверхности пораженных органов плодовые тела видны невооруженным глазом, как небольшие точки. В начале развития – белые, затем желтые, при созревании – темно-коричневые или черные. Окраска связана со зрелостью наружного слоя – перидия. От наружных клеток перидия отходят различные отроски или аппендиксы (придатки). Строение придатков различно. Они состоят из мертвых клеток с сильно утолщенными стенками. Придатки бывают простыми, гифоподобными, дихотомически разветвленными, с луковицеобразным расширением, спирально закрученными. Они отличаются по степени окраски и расположению на плодовом теле.
Аски – мешковидные, веерообразно располагаются в полости клейстотеция. У различных родов количество асков в клейстотеции разное. У Sphaerotheca, Podosphaera – по одной, у родов Erysiphe, Microsphaera, Uncinula, Leveillula, Phyllactinia – несколько или много асков. Число асков в клейстотеции у некоторых видов рода Leveillula может достигать 40.
Биология
Представители класса Гифомицеты относятся к различным экологическим группам грибов: почвенным, ксилофильным, паразитическим, хищным, водным микрофильным, энтомофильным.
Размножаются Гифомицеты, как и другие несовершенные грибы класс Целомицеты (Coelomycetes), Агономицеты (Agonomycetes), вегетативно и бесполыми спорами.
Вегетативное размножение может осуществляться неспециализированными участками мицелия, дающими начало новому мицелию. Отмечается, что мицелий грибов обладает высокой способностью к регенерации. К специализированным структурам вегетативного размножения относят так же хламидиоспоры. Последние, способны сохраняться в неблагоприятных для развития вида условиях.



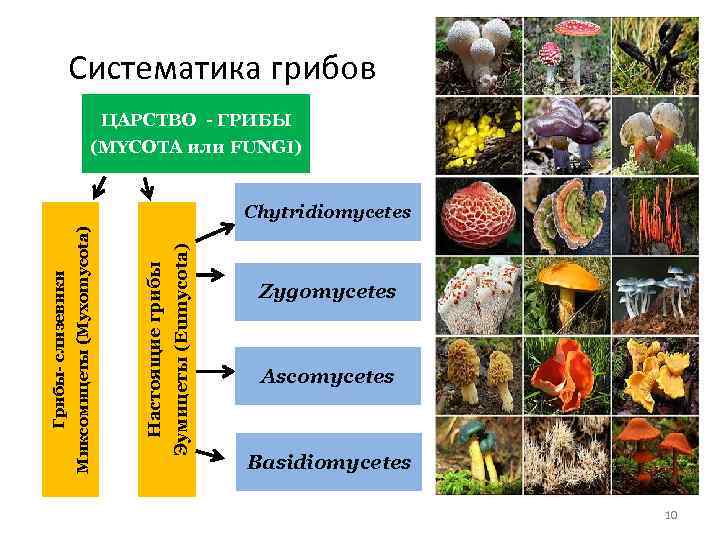
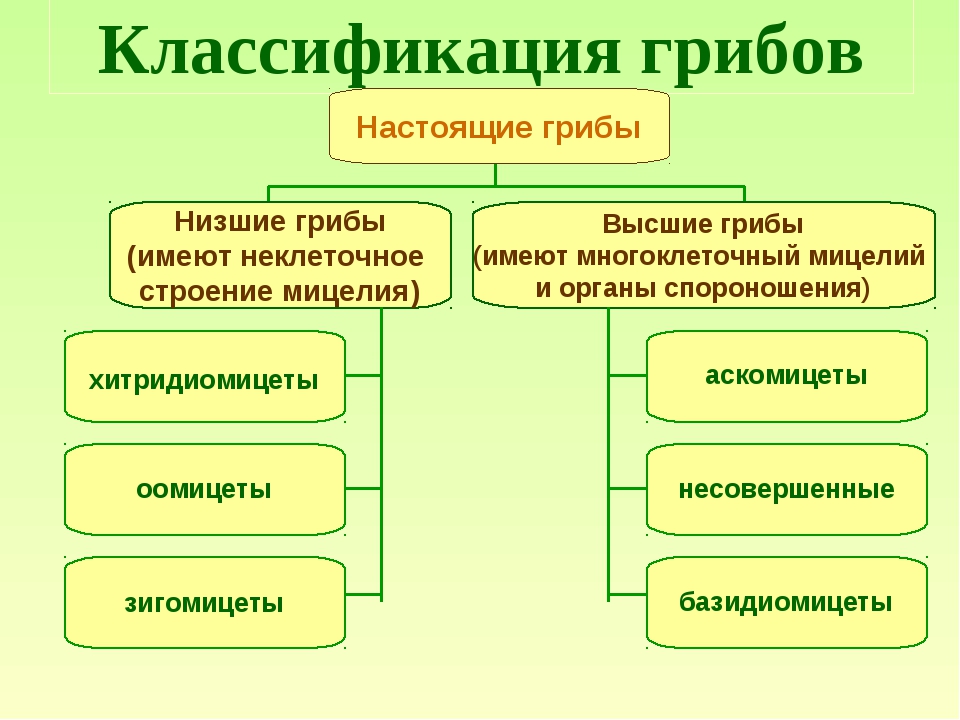
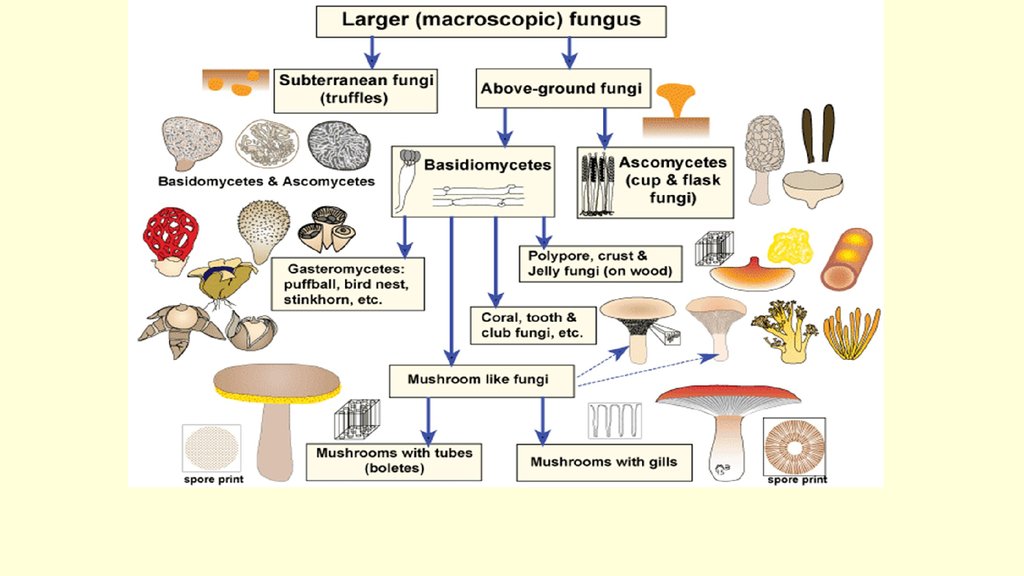
Место класса Гифомицеты в макросистеме грибов
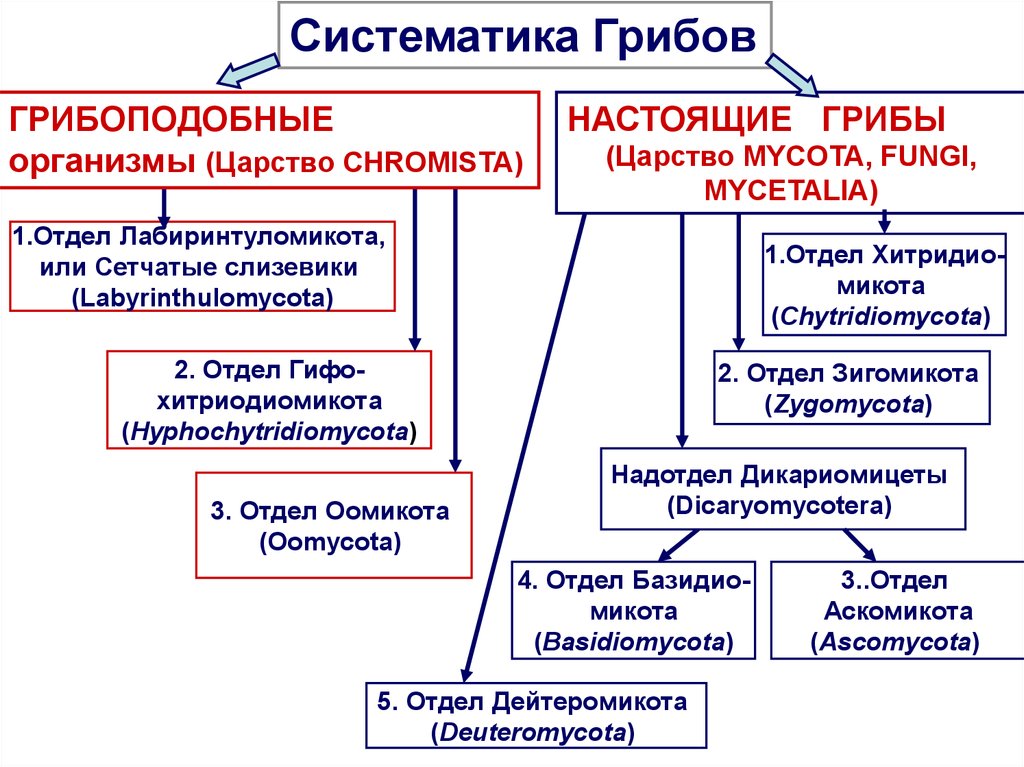
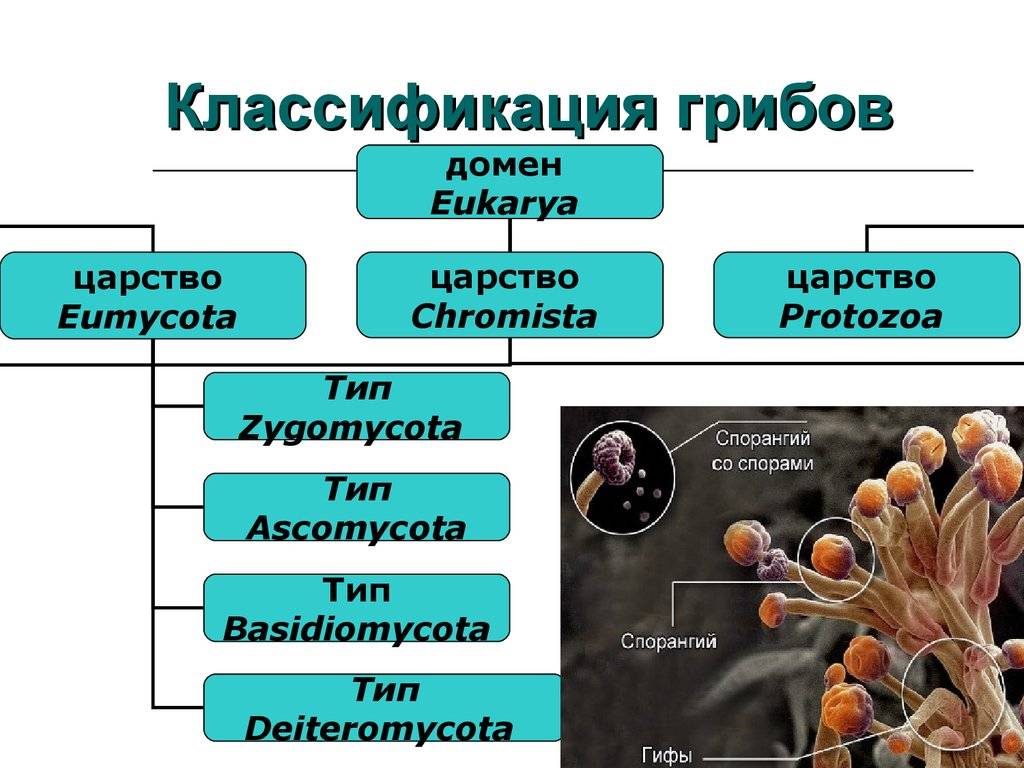
Класс Гифомицеты (Hyphomycetes) вместе с ещё двумя классами Целомицеты (Coelomycetes), Агономицеты (Agonomycetes) относят типу Anamorphic fungi, включенный в царство Настоящих грибов Fungi, Mycota .
В класс Гифомицеты Hyphomycetes включен только один порядок Гифомицеты (Hyphomycetales).
Представители класса Гифомицеты, как и все Несовершенные грибы, утратили в ходе эволюции половую стадию (телеоморфа) и размножаются бесполым путем – вегетативно и конидиями. Виды этой группы связывают по происхождению с Аскомицетами и Базидиомикотами и рассматриваются в качестве новой, находящейся в процессе становления эволюционной ветви грибов полифилетического происхождения. Эволюция группы на данном этапе направлена на усовершенствование конидиального аппарата (конидии, конидиеносцы), как основного органа размножения и компенсацию отсутствия полового процесса прочими механизмами, обеспечивающими этим грибам новые механизмы микроэволюции.
Выявление принадлежности этой группы к типам Аскомицетов или Базидиомикот грибов происходит только исходя из молекулярно-генетических и микроморфологических признаков.
У многих видов уже установлена связь между анаморфами (видами с бесполым размножением) и телеоморфами (видами с половым размножением). На этом основании многие исследователи рекомендуют относить виды с известными телеоморфами к соответствующим группам аскомицетов или базидиомикот. Однако, прагматический подход приводит к правомерности существования формальной, построенной исключительно на морфологических признаках системе, далекой от филогенетической, но пригодной для идентификации,системе, принятой в большинстве случаев в настоящее время.