Вред, наносимый тафринами
Эти грибы являются облигатными паразитами высших растений. Они вызывают заболевания многих растений, используемых в сельском хозяйстве. Тафрины вызывают такие болезни садовых деревьев, как «курчавость листьев», «ведьмины метлы», «дутые плоды» и «кармашки». От них страдают персики, сливы, вишня, миндаль, груша и другие плодовые деревья.
Такие заболевания становятся причиной снижения урожая. Деревья ослабляются, становятся подверженными вторичным заболеваниям, при сильном поражении растение может погибнуть. У больных деревьев в значительной мере снижается урожайность. Заболевания, вызываемые тафриной, поражают и дикорастущие деревья и кустарники, например, черемуху и терн. Кроме того, от них могу страдать клен, береза, ольха, вяз, дуб, тополь. Появление таких грибов в городских насаждениях снижает их декоративность.
Симптомы заражения растений тафринами
Внешне различные виды тафрины сложно отличить друг от друга, поэтому в систематике определения видов грибов важное значение имеет характер повреждений растения-хозяина. Заражение этими грибами может вызывать у растений различные уродства – патологические разрастания листьев, деформация разных частей, недоразвитость семян и тому подобное
Такие нарушения связаны с тем, что грибы производят вещества, обладающие гормональной активностью, например, гетероауксин и цитокинины.
Если мицелий развивается под кутикулой растения, то чаще всего деформации не происходит, лишь образуются на поверхности пятна различного размера. Пятнистость формируется из-за некроза тканей. Если происходит гипертрофия клеток, то на верхней стороне поврежденной части растения возникают утолщения, а в нижней, напротив, появляются углубления.
Если заражению подвергаются побеги, то стимулируется рост почек боковых побегов, в результате появляются «ведьмины метла», длина которых может доходить до 3-х метров. Такие метла состоят из тонких и коротких побегов, нередко сильно разветвленных. Больные побеги почти не плодоносят, их листья неправильной формы, мелкие, быстро опадающие. Еще один симптом заражения грибами тафринами – утолщение и искривление стеблей растения.
Состав
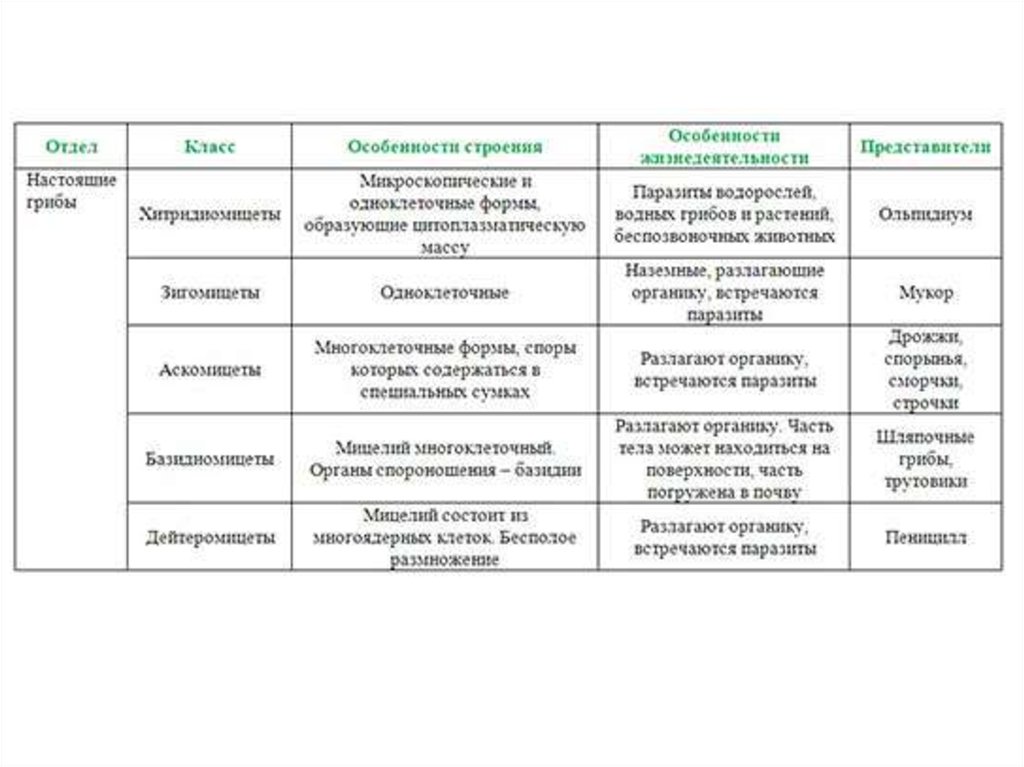
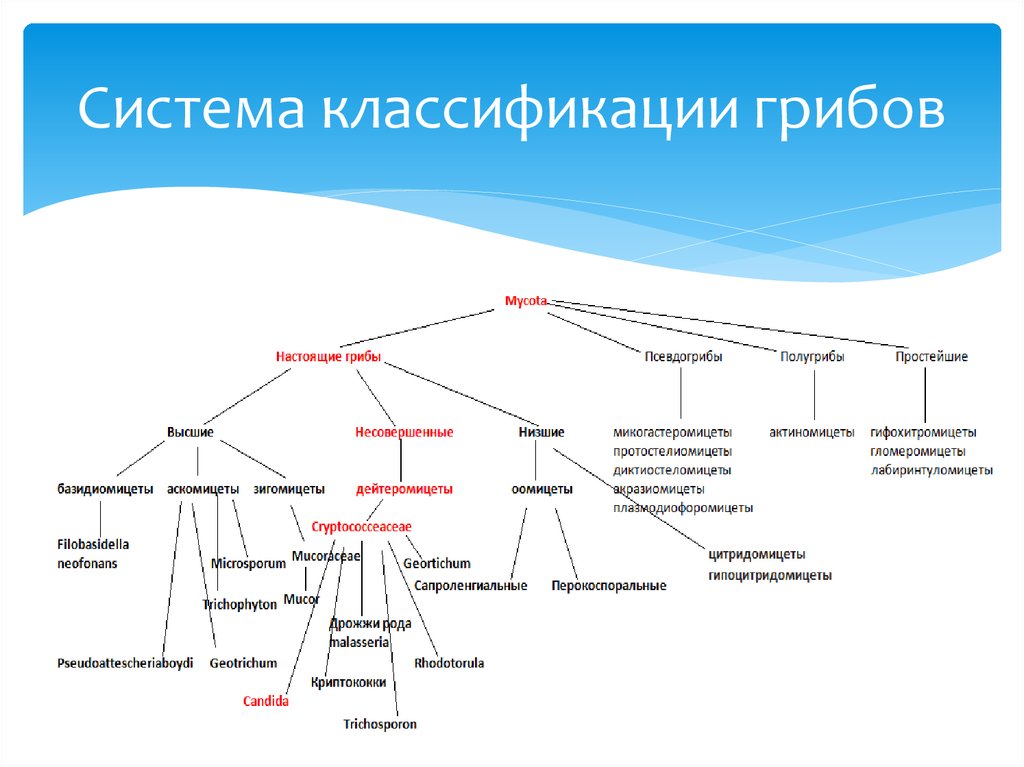
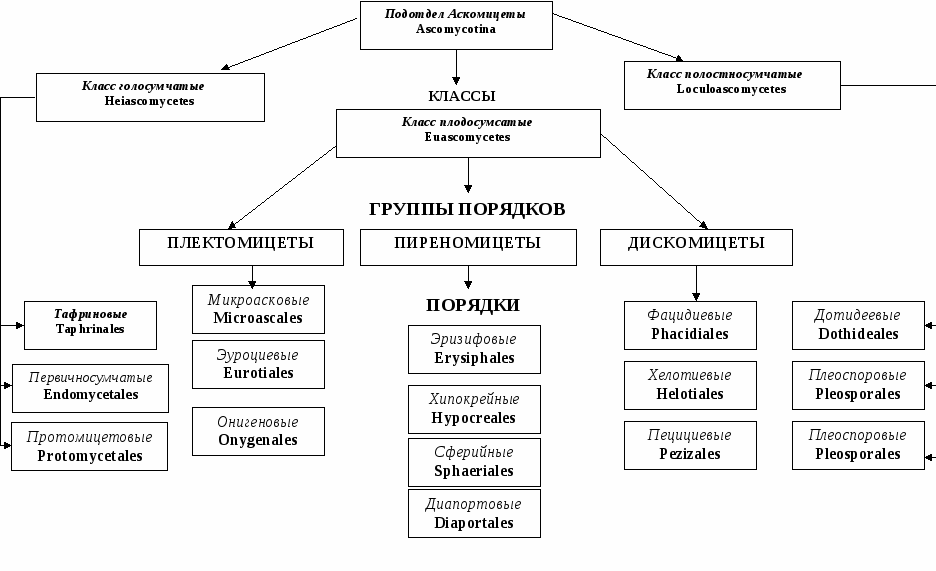
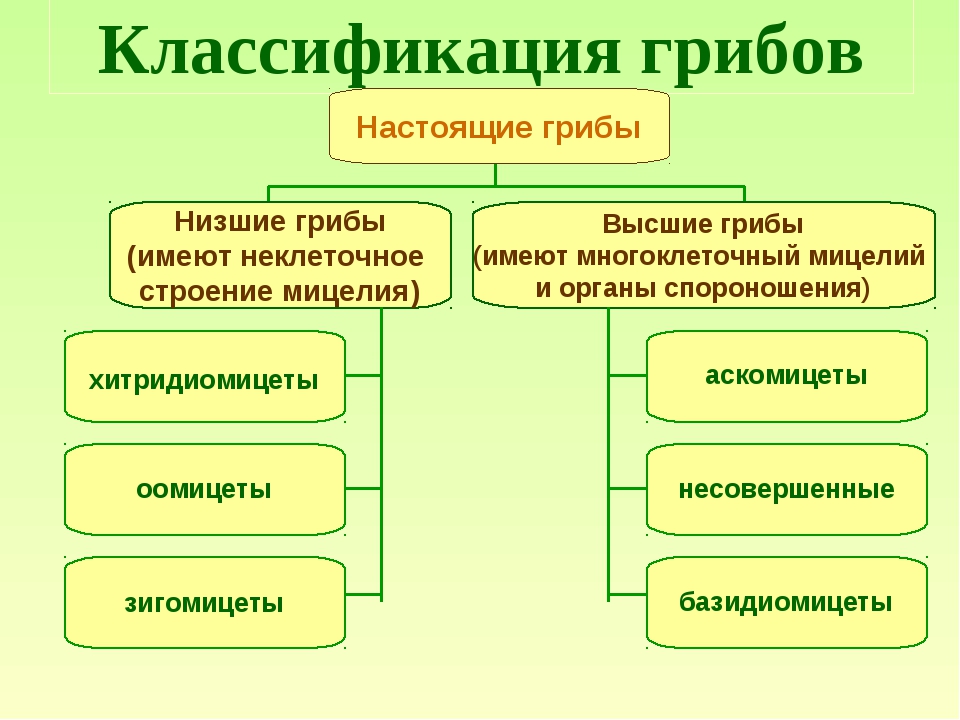
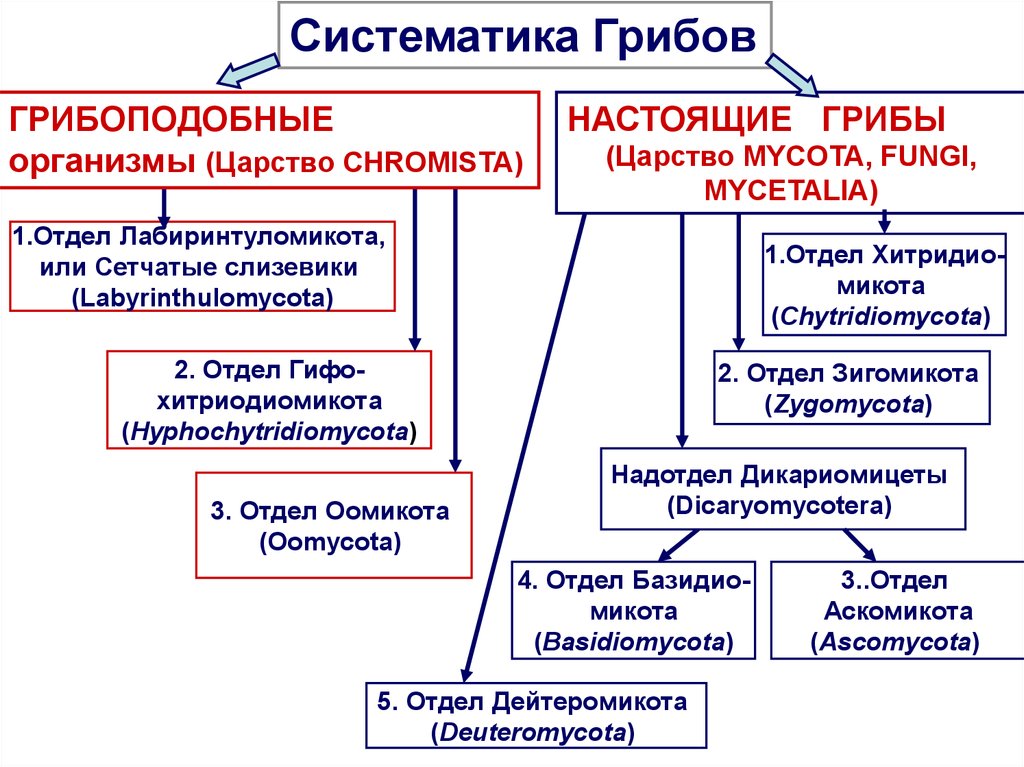
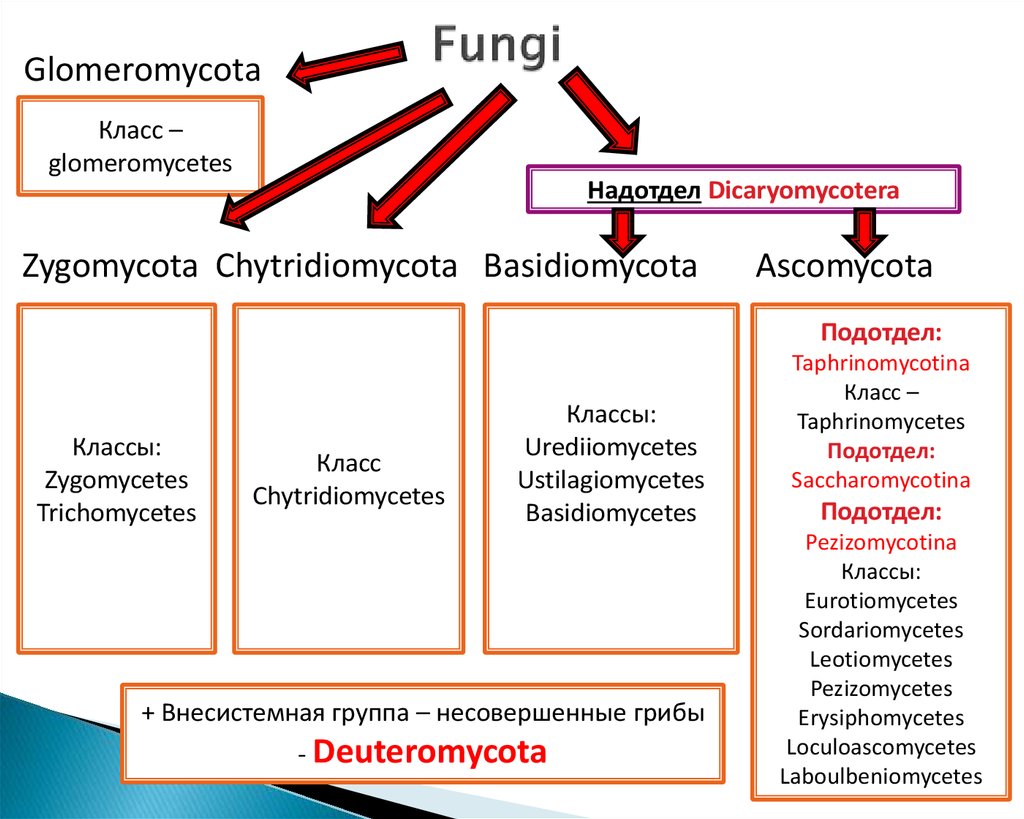
Таксон включает 4 класса.
- Неолектомицеты (Neolectomycetes) — грибы, растущие в ассоциациях с деревьями и образующие яркоокрашенные плодовые тела размерами до 7 см. Характер взаимоотношений их с растениями ещё не выяснен.
- Pneumocystidomycetes — дрожжевые грибки, патогенные для человека и других млекопитающих (известны, в частности, как возбудители пневмоцистной пневмонии).
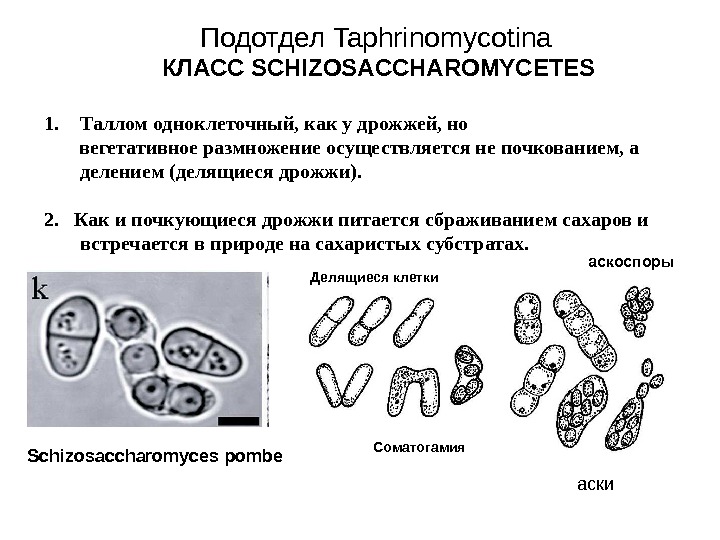
- Схизосахаромицеты (Schizosaccharomycetes) — делящиеся дрожжи, вызывающие спиртовое брожение сахаристых жидкостей и используемые человеком для приготовления напитков брожения (включая некоторые сорта пива).
- Тафриномицеты (Taphrinomycetes) — паразиты высших растений, вызывающие пятнистость листьев и стеблей, поражения плодов, ведьмины мётлы и другие заболевания.
В системах, существовавших до конца XX века, представители тафриномицетов не объединялись в один таксон. Neolecta, единственный род класса Неолектомицеты, первоначально был описан как представитель рода Geoglossum, ныне входящий в порядок Leotiales. Семейства, вошедшие в класс Taphrinomycetes, относили к подклассу голосумчатых, или гемиаскомицетов (Hemiascomycetidae), в который входил также порядок эндомицетовых (Endomycetales, приблизительно соответствует современному классу Сахаромицеты), к которому относили делящиеся дрожжи. Организмы из современного класса Pneumocystidomycetes были открыты в 1912 году и описаны как простейшие (трипаносомы), а к царству грибов отнесены только в конце XX века на основании молекулярно-филогенетического анализа.
Филогения
Согласно данным традиционного кладистического анализа, подотдел представляет собой монофилетическую группу, однако между классами существуют значительные различия в рибосомных генах. Было высказано предположение о парафилетичности подотдела, которое нашло определённое подтверждение в данных молекулярно-филогенетических исследований, проводившимися в 2000 году и соавторами и в 2001 году У. Эрикссоном. Последующие исследования, однако, вновь подтвердили монофилию подотдела Taphrinomycotina как базальной группы в отделе Ascomycota, причём филогенетические связи между классами, входящими в подотдел, представляются следующей кладограммой:
| Taphrinomycotina |
| ||||||||||||||||||
В 2011 году состав подотдела Taphrinomycotina пополнился ещё одним классом: Archaeorhizomycetes (археоризомицеты), в который был включён единственный род Archaeorhizomyces. Согласно предварительным результатам филогенетического анализа, данный класс представляет собой сестринскую группу для клады, образованной схизосахаромицетами и пневмоцистидомицетами. Позднее появились данные, заставляющие вновь сомневаться в монофилии Taphrinomycotina; так, в исследовании 2017 года основная часть Taphrinomycotina, Neolectomycetes и Archaeorhizomycetes представлены как три последовательно обособляющиеся клады в основании отдела Ascomycota.
Статья составлена с использованием следующих материалов:
Пересыпкин В.Ф. Болезни сельскохозяйственных культур. Том 3. Болезни овощных и плодовых культур, Киев: Урожай, 1991. — 208 с.
2.
Пересыпкин В.Ф. Сельскохозяйственная фитопатология, 4-е изд., перераб. и доп. – Москва: Агропромиздат, 1989. – 480 с.




3.
Пидопличко Н.М. Грибы – паразиты культурных растений. Определитель. Том 1. Грибы совершенные, К.: “Наукова думка” 1977. – 296 с.
4.
Поликсенова В.Д., ХрамцовА.К., Пискун С.Г., Методические указания к занятиям спецпрактикума по разделу: «Микология. Методы экспериментального изучения микроскопических грибов» для студентов 4 курса дневного отделения специальности «G 31 01 01 – Биология» – Мн.: БГУ, 2004 – 36 с.  Скачать >>>
5.
Станчева Й. Атлас болезней сельскохозяйственных культур. Том 2. Болезни плодовых, ягодных, орехоплодных культур и винограда, София –Москва, изд. «Пенсофт»,: 2005. — 196 с.
Изображения (переработаны):
6.
Ванек Г., Корчагин В.Н., Тер-Симонян Л.Г. Атлас болезней и вредителей плодовых, ягодных, овощных культур и винограда. Братислава.–М. Природа. Агропромиздат. 1989 г. 413 с., илл. Иллюстрации из книги.
7.
Ascus and ascospores (Taphrina deformans), by Gerald Holmes, California Polytechnic State University at San Luis Obispo, Bugwood.org, по лицензии CC BY-NC
Свернуть
Список всех источников
Жизненный цикл и онтогенез
Ядерный жизненный цикл — гаплоидно-дикариотический, преобладает дикариотическая фаза.
Гаплоидные аскоспоры прорастают бластоспорами, у некоторых видов прорастание аскоспор происходит ещё до освобождения из сумок. Бластоспоры дают начало анаморфной гаплоидной дрожжевой фазе, состоящей из почкующихся клеток, питающихся сапротрофно. Тафриновые дрожжи описывают в составе анаморфного рода Lalaria, введённого в 1990 году. Заканчивается гаплоидная фаза копуляцией (слиянием) клеток и образованием дикариона. У некоторых видов дикариотизация может происходить без спаривания клеток, путём деления ядра. При искусственном разведении дикариотический мицелий обычно не развивается, зато отмечено появление особых толстостенных клеток. Предполагают, что эти клетки способны выживать при неблагоприятных условиях, то есть в природе выполняют функцию хламидоспор. Дикариотический мицелий, в отличие от большинства других аскомицетов (кроме сахаромицетов), питается самостоятельно, а не за счёт гаплоидной фазы. Дикариофаза ведёт исключительно паразитический (биотрофный) образ жизни, развиваясь между клетками заражённого растения.
Схема аскогенеза Taphrina-типа:
Для рода характерен специфический способ формирования сумок, названный Taphrina-типом. Под кутикулой растения образуется слой клеток, называемых аскогенными клетками, которые представляют собой фрагменты недифференцированных гиф. В них происходит слияние ядер (кариогамия), диплоидные клетки увеличиваются в размерах и их ядра делятся митозом. Одновременно с митозом аскогенная клетка дифференцируется на базальную и предасковую (проаск) клетки, каждая из них получает одно дочернее диплоидное ядро. Затем в предасковой клетке осуществляется мейоз, и формируется короткая 8- или 4-споровая сумка, после спорообразования базальная клетка дегенерирует.

Базальные клетки формируются не у всех видов; также сумки, имеющие собственные оболочки, могут формироваться внутри аскогенных клеток, стенки последних затем прорываются наружу. Образование аскоспор тоже имеет некоторые особенности, отличающие тафрину от других аскомицетов. Каждое из гаплоидных ядер с участками цитоплазмы окружается плазмалеммой, впячивающейся внутрь сумки от её стенки, затем мембрана формирует споровую стенку; независимый от плазмалеммы мембранный мешок отсутствует. После созревания спор верхушка сумки вскрывается простой щелью, и споры освобождаются пассивно или выбрасываются с силой.
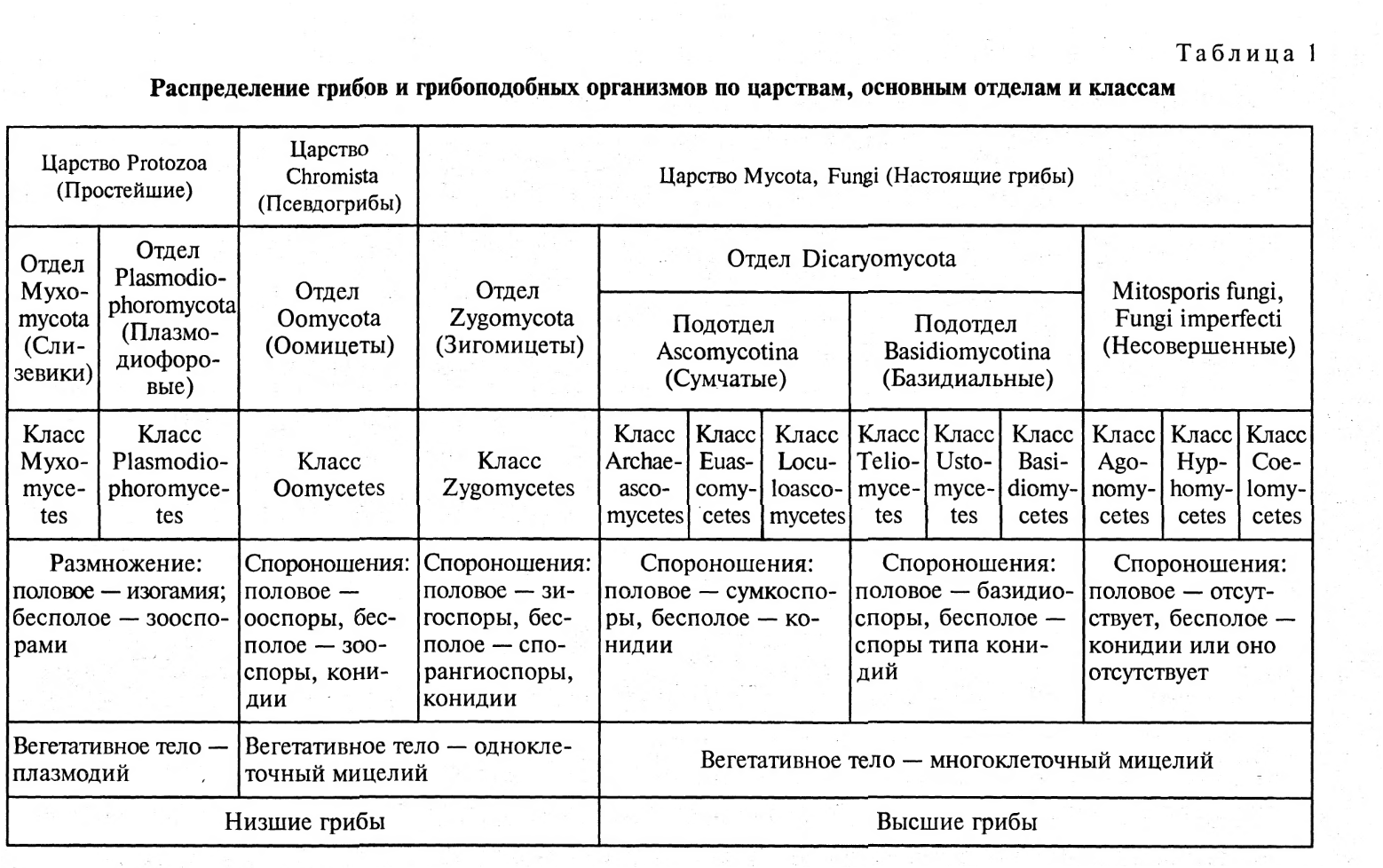
Голосумчатые (Hemiascomyeetidae)
Сумки располагаются свободно, плодовые тела отсутствуют. Среди представителей голосумчатых грибов фитопатогены встречаются в двух порядках.
Порядок Endomycetales (первичносумчатые, или сумчатые, дрожжи). Сумки одиночные, почкующиеся или расположенные на выростах мицелия. Представители этого порядка часто способствуют патологическому слизетечению, развиваясь в вытекающем из трещин коры соке.
Endomyces vernalis вызывает млечное слизетечение у лиственных пород. Меры борьбы: своевременное залечивание травм, фитосанитария.
Порядок Taphrinales(тафриновые). Представители этого порядка — облигатные паразиты с узкой специализацией. Вызывают деформации различных органов древесных растений. Поверхность пораженных органов покрывается рыхлым слоем сумчатого спороношения гриба, который быстро исчезает.
Taphrina acerinum вызывает компактное ветвление побегов клена.
Taphrina aurea — возбудитель деформации листьев тополя.
Taphrina alni паразитирует на чешуйках сережек ольхи, вызывает деформацию листьев ольхи.
Taphrina pruni является причиной деформации плодов и листьев косточковых пород («кармашки плодов»), в том числе черемухи и персика.
Taphrina betulina вызывает компактное ветвление побегов березы («ведьмина метла»).
В качестве мер борьбы используют методы фитосанитарии и химической защиты, применяемые в основном в садоводстве.
Характеристика
Представители подотдела разнообразны по морфологии и жизненным циклам. Они могут существовать в мицелиальной или дрожжевой форме, для многих видов характерен мицелиально-дрожжевой диморфизм. Ядерный жизненный цикл гапло-дикариотический или гапло-диплоидный, у некоторых представителей может быть гаплоидный. Клеточные стенки содержат очень незначительное количество хитина, а иногда и вовсе не содержат. Плодовые тела, кроме представителей класса неолектомицетов, отсутствуют, у мицелиальных представителей сумки формируются непосредственно на мицелии, не образующем аскогенных гиф; у некоторых видов половой процесс неизвестен. Верхушки сумок не содержат структур, связанных с активным выбрасыванием спор, но споры могут отделяться и активным способом, причём активное или пассивное их отделение может наблюдаться у одного и того же вида в зависимости от внешних условий. Крючки при формировании сумок у тафриномицетов также отсутствуют. Сумки могут формировать плотный слой, подобный гимению, но не содержащий стерильных элементов.
Филогения
Согласно данным традиционного кладистического анализа, подотдел представляет собой монофилетическую группу, однако между классами существуют значительные различия в рибосомных генах. Было высказано предположение о парафилетичности подотдела, которое нашло определённое подтверждение в данных молекулярно-филогенетических исследований, проводившимися в 2000 году и соавторами и в 2001 году У. Эрикссоном. Последующие исследования, однако, вновь подтвердили монофилию подотдела Taphrinomycotina как базальной группы в отделе Ascomycota, причём филогенетические связи между классами, входящими в подотдел, представляются следующей кладограммой:
| Taphrinomycotina |
| ||||||||||||||||||
В 2011 году состав подотдела Taphrinomycotina пополнился ещё одним классом: Archaeorhizomycetes (археоризомицеты), в который был включён единственный род Archaeorhizomyces. Согласно предварительным результатам филогенетического анализа, данный класс представляет собой сестринскую группу для клады, образованной схизосахаромицетами и пневмоцистидомицетами. Позднее появились данные, заставляющие вновь сомневаться в монофилии Taphrinomycotina; так, в исследовании 2017 года основная часть Taphrinomycotina, Neolectomycetes и Archaeorhizomycetes представлены как три последовательно обособляющиеся клады в основании отдела Ascomycota.