Меры борьбы
Профилактические мероприятия
(сокращение и ликвидация мест выплода комаров):
- Осушение болот.
- Уничтожение ненужных водоемов.
- Устранение утечек воды.
- Укрепление берегов водоемов.
- Санитарный надзор при проектировании, строительстве, эксплуатации водохранилищ, оросительных и дренажных систем, рисовых чеков.
- Плотное закрывание бочек, кадок, цистерн и других емкостей с водой летом.
- Засетчивание окон и вентиляционных отверстий.
Истребительные мероприятия
(проводятся по показаниям):
- Противоличиночные обработки инсектицидами.
- Противоимангинальные обработки инсектицидами.
Ларвициды. Все анофелогенные водоемы в населенном пункте подлежат обработке бактериальными инсектицидами. Обработка в летнее время проводится один раз в 10 дней. Применение на водоемах химических интоктицидов нежелательно. Разрешенные бактериальные препараты – бактицид, ларвиоль-паста.
Химические средства. Основной способ уничтожения эндофильных комаров – внутренняя обработка помещений инсектицидами длительного действия.
Экзофильные виды комаров уничтожают путем обработки дневок в растительности вблизи мест выплода, а при необходимости и помещений, куда комары могут залетать. Растительность вокруг мест выплода обрабатывают инсектицидами в радиусе 50–100 метров.
Морфология
Имаго. Комар размером 6–8 мм. Морфология вида подвержена значительным колебаниям по окраске и величине. Экземпляры, обитающие на юге, характеризуются более светлой окраской и меньшими размерами.
Голова с боков усажена торчащими темно-бурыми чешуйками, сверху расположен направленный вперед пучок длинных беловатых чешуек и волосков. Усики бурые. Щупики и хоботок бурые, преимущественно в прилегающих чешуйках.
Среднеспинка в средней части с широкой, кпереди суживающейся продольной полосой сероватого цвета. На передней половине данной полосы расположены, как правило, 2–3 неясные буроватые полоски. По переднему краю среднеспинки у головы располагается пучок беловатых чешуек. Боковые отделы в передней части бурые, в задней – черно-бурые. Волосяной покров в средней части и на щитке светло-бурый, в остальной части, особенно над основаниями крыльев, темно-бурый.
Ноги бурые, с нижней стороны бедер и внутренней стороны голеней более светлые, лапки темные.
Крылья покрыты чешуйками буроватого оттенка, с черными пятнами более темных черно-бурых чешуек, расположенных при основании радиальной и медиальной вилок. Чешуйки по краю крыла и на его вершине светло-желтые.
Жужжальца желтые, с буроватой головкой.
Брюшко бурое или черно-бурое, часто с более светлыми пятнами на боковых отделах тергитов, покрыто довольно длинными золотисто-желтыми или буроватыми волосками.
Половой диморфизм. Разнополые особи отличаются строением половых органов.





Самка. Хоботок состоит из верней и нижней губы, подглоточника, пары верхних и пары нижних челюстей. Щупальца длинные, примерно равны хоботку.
Самец. Парные части хоботка редуцированы. Щупальца также же длинные, но с явственно утолщенными конечными члениками.
Яйцо оборудовано плавательной камерой, занимающей среднюю треть яйца, и оторочкой. Структура и окраска экзохориона различается, но характерна для отдельных подвидов. Она зависит от местных изменений строения наружной оболочки и состоит из светлых и темных пятен и полос.
Личинка IV стадии. По причине большого биологического разнообразия мест обитания и условий развития личинки вида изменчивы по внешнему виду, величине и окраске. Особи северных широт отличаются более крупными размерами и более темной пигментацией на хитиновых образованиях (голове, стигмальных пластинках и пр.).
Общая окраска тела варьирует от серо-желто-зеленой до черно-зеленой.
Пигментация головы изменчива – от светло-коричневых пятен до черно-коричневых полос на общем темном фоне.
Усики почти прямые, до 2/3 длины головы, с более темным концом. Волосок короткий, в среднем равен ширине усика у места своего отхождения, лежит у основания и состоит из 4–6 тонких веточек.
Стигмальная пластинка крупная. Твердые хитиновые образования сильно пигментированы. Передняя лопасть с темным основанием, остальная часть заметно светлее. Стигмы на некотором расстоянии от центральной пластинки отростков не имеют.
Гребень на боковых пластинках VIII членика состоит из 6–10 (чаще 7–9) крупных шиповидных зубцов, между ними находятся от 1 до 4 более мелких зубцов.
Плавник составлен из 18–20 пучков. Жабры в среднем равны длине седла, обе пары одной длины.
Куколка, как и у всех представителей рода Anopheles, сбоку имеет вид запятой, сверху – неправильного овала. Тело состоит из двух отделов: массивной головогруди и тонкого уплощенного дорсовентрального брюшка. Глаза сильно пигментированы и просвечивают через прозрачную кутикулу. Над ними по бокам спинки отходят куколочные рожки, суженные у основания и сильно расширяющиеся к свободному концу. Их отверстия скошены внутрь и при нахождении куколок на поверхности лежат в плоскости поверхностной пленки. От основания рожек отходит трахея, соединяющая полость рожек с переднегрудной стигмой развивающегося комара.
Брюшко куколки состоит из девяти члеников. На заднем крае спинной стороны VIII членика находятся два плавника, заходящие друг за друга внутренними краями. Каждый плавник имеет вид неправильной овальной пластинки, сидящей на суженном основании. Пластинка укреплена продольным срединным ребром, не доходящим до ее свободного края. От конца ребра отходит волосок, у его основания располагается более короткий добавочный волосок.
Разновидность
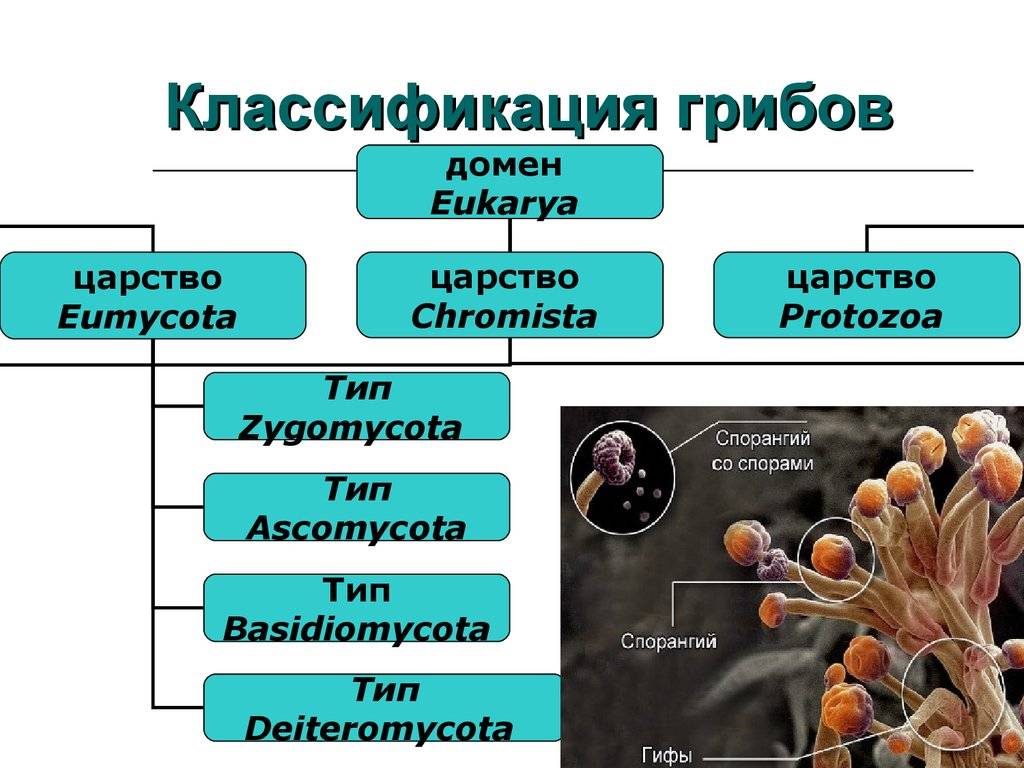
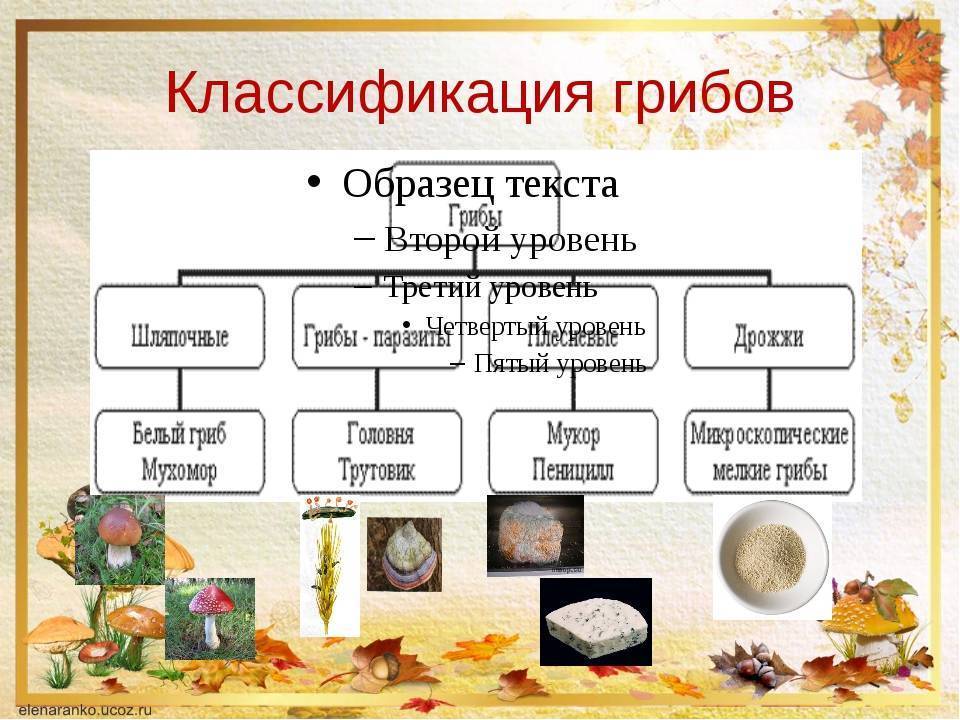
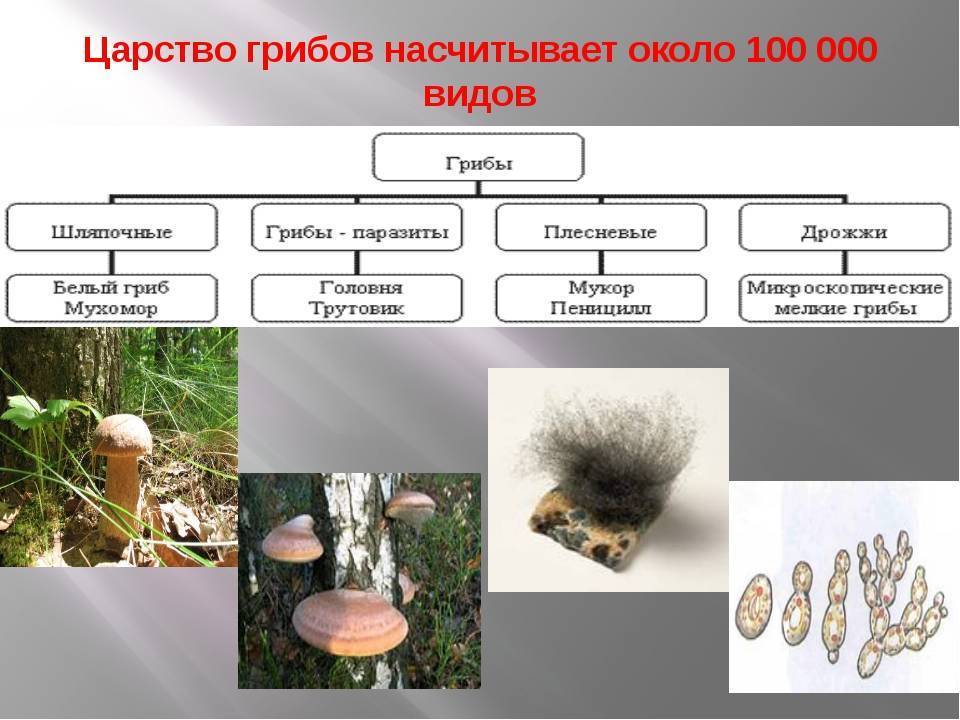
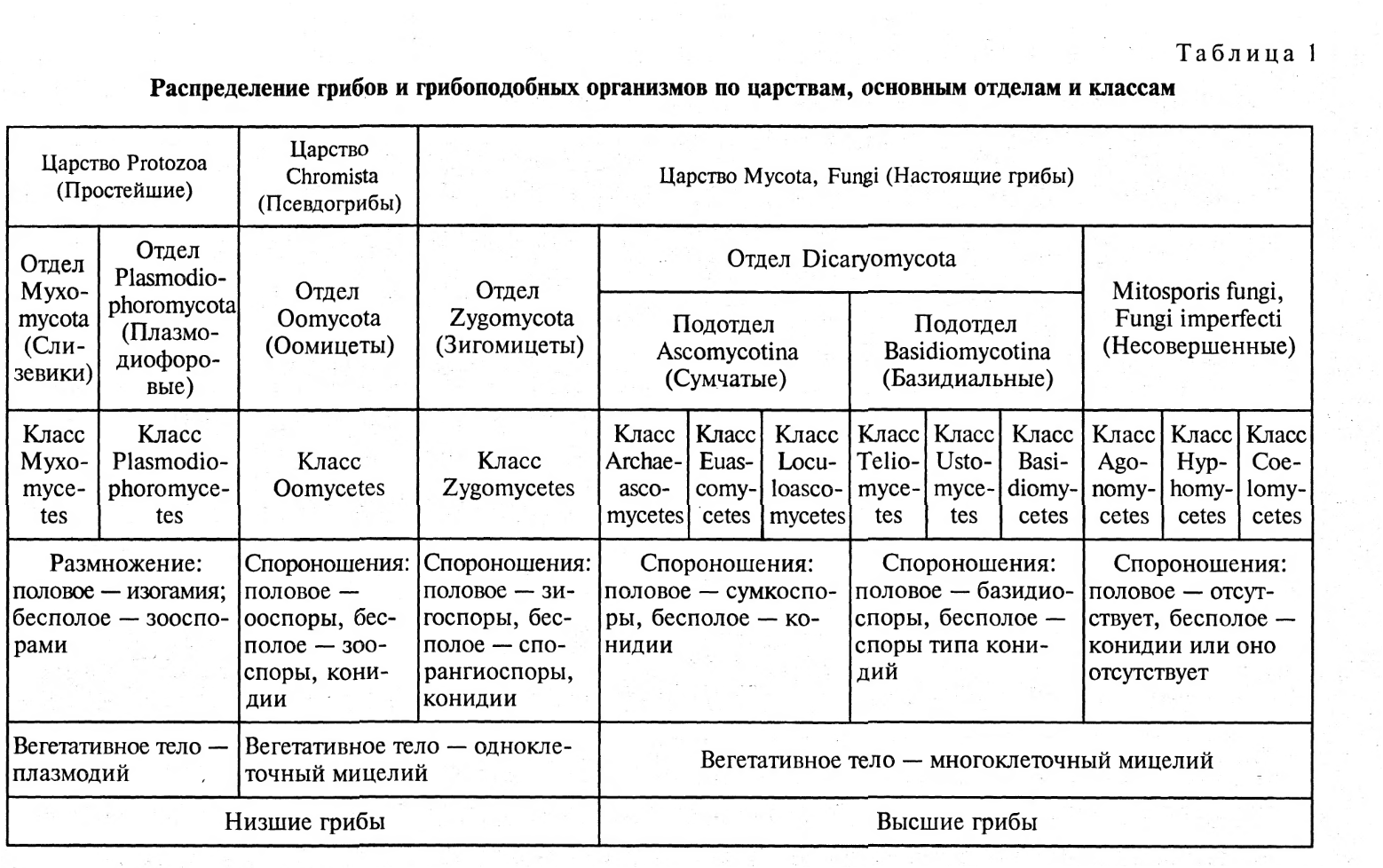
По состоянию на июнь 2017 года Index Fungorum принимает пять видов Amyloporia :
- A. nothofaginea Rajchenb. & Gorjón (2011) – южные Анды
- A. sinuosa (фр.) Rajchenb., Gorjón & Pildain (2011)
- A. stratosa (JEWright & JRDeschamps) Rajchenb., Gorjón & Pildain (2011) – Южная Америка, Тасмания
- A. subxantha (YCDai и XSHe), BKCui и YCDai (2013 г.) – Китай
- A. turkestanica (Pilát) Бондарцев (1953) – Средняя Азия
Index Fungorum показывает 30 таксонов, связанных с родовым названием Amyloporia . Несколько видов, когда-то помещенных в этот род, с тех пор были перенесены в другие роды:
- Amyloporia alpina (Litsch.) Domański (1974) = Antrodia alpina (Litsch.) Gilb. И Риварден (1985)
- Amyloporia calcea (фр.) Бондарцев и Зингер (1944) = Antrodia calceus (фр.) Тейшейра (1992)
- Amyloporia carbonica (Overh.) Vampola & Pouzar (1993) = Antrodia carbonica (Overh.) Ryvarden & Gilb. 1984)
- Amyloporia crassa (P.Karst.) Bondartsev & Singer (1941) = Antrodia crassa (P.Karst.) Ryvarden (1973)
- Amyloporia lenis (P.Karst.) Bondartsev & Singer (1941) = Sidera lenis (P.Karst.) Miettinen (2011)
- Amyloporia pinea B.K.Cui & YCDai (2013) = Antrodia pinea (BKCui & YCDai) В. Спирин (2015)
- Amyloporia sitchensis (DVBaxter) Vampola & Pouzar (1993) = Antrodia sitchensis (DVBaxter) Gilb. И Риварден (1985)
- Amyloporia sordida (Ryvarden & Gilb.) Vampola & Pouzar (1993) = Antrodia sordida Ryvarden & Gilb. (1984)
- Amyloporia xantha (Fr.) Bondartsev & Singer (1941) = Antrodia xantha (Fr.) Ryvarden (1973)
Развитие
Имаго. Зимует во взрослой фазе оплодотворенной самки как эндофильный вид в различных нежилых помещениях, часто совместно с сельскохозяйственными животными. В природе – в пещерах, норах. В северной и восточной частях ареала комары выносят многомесячное воздействие очень низких температур. Температурный минимум развития составляет +7–10 °С.За теплое время года вид дает 2 поколения на севере и 4–6 на юге. Максимум численности наблюдается в середине лета, при очень жаркой погоде отмечается снижение численности.




Главная масса комаров концентрируется в населенных пунктах или недалеко от них. Питаются в основном кровью сельскохозяйственных животных, домашних и диких птиц, в меньшей степени людей, но при отсутствии животных основной источник кровососания – люди. Питание кровью необходимо самкам. Автогенного развития яиц у данного вида не наблюдается. Для данного вида характерна гонотрофическая гармония – одного полного насыщения кровью достаточно для развития и откладки яиц.
Период спаривания. Копуляция проходит на лету при роении. Яйца в теле самок при температуре +7,5°С созревают в течение 23 суток, при +1 °С – 7 суток, а при +20°С – 4 суток. При оптимальной температуре +25–30°С – в течение 2–3 суток. После созревания яиц самки вылетают с мест дневок и ищут водоемы для откладывания яиц. Самки откладывают яйца поодиночке, впоследствии те могут сцепляться друг с другом, но не склеиваться. Количество яиц колеблется от 100 до 500 штук.
Яйцо. Как у всех кровососущих комаров, длительность эмбрионального развития может колебаться от 40 часов до 8 суток, при температуре +23–24°С – 3–6 суток.
Личинка. Места обитания – разнообразные по типу, глубине и величине водоемы, хорошо освещенные солнцем на севере и затененные на юге. В южных районах обитают в слабо солоноватой воде. Оптимальная скорость развития наблюдается при температуре +29–30°С – 5–7 суток, при +15–16°С – до 30 суток.
Куколка. Зрелая личинка IVстадии после линьки переходит в куколку. Куколки не питаются и с внешней средой связаны только процессами газообмена. В нормальном состоянии они свешиваются вниз с поверхностной пленки воды. Куколки способны к быстрым активным движениям и опускаются на дно водоема при внешних раздражениях. В спокойном состоянии куколки долго находятся на поверхности воды. Длительность куколочной фазы в среднем не превышает 2–2,5 суток. Первыми всегда вылетают самцы.
Морфологически близкие виды
По внешнему виду (морфологии) имаго близок Комар малярийный камышовый (Anopheles hyrcanus). Отличается наличием на переднем крае крыльев светлых чешуек, образующих два пятна, и торчащими чешуйками щупиков.
Кроме того, отмечается, что вид Anopheles maculipennis разделяется на несколько рас, иногда они трактуются как подвиды. Шесть из них представлены в фауне Палеарктики: maculipennis, messefe, melanoon (subalpinus), labranchiae, atrorparvus, sacharovi.
Высказываются предложения о возведении названных рас в ранг видов. Однако такая крайняя точка зрения общего признания не получила, поскольку формы комплекса maculipennisближе друг к другу, чем к другим формам. Намечающиеся группы внутри комплекса иногда предлагается рассматривать в качестве видов. Такая трактовка принята многими, в частности авторами всесветного каталога комаров. В составе фауны Палеарктики вместо одного вида Anopheles maculipennis признаются три: maculipennis с подвидами maculipennis, messefe, melanoon; labranchiae с подвидами labranchiae и atrorparvus; и sacharovi.
Североамериканский Аn. occidentalis тоже рассматривается в качестве отдельного вида. Данные четыре вида совместно с некоторыми другими близкими и образуют группу maculipennis, которая входит в состав одной из шести групп видов, на которые подразделяется подрод Anopheles.
Сторонники дробления комплекса maculipennis на большее или меньшее количество видов указывают на различие в дорсальной поверхности яиц и наличие биологических отличий между формами.
Однако не обнаружено никаких достоверных различий между членами комплекса ни по личинкам, ни по имаго, ни по строению гипопигия (основным принципам классификации комаров). Известны только некоторые отличия sacharovi, но они несущественны и имеют характер отличий только подвидового ранга.
Что касается биологических особенностей, отмечается, что довольно существенные внутривидовые различия выявляются у многих других видов комаров. В этом случае имеют дело, скорее всего, с биотипами. Поэтому вопрос о дроблении как maculipennis, так и других видов, признать оправданным нельзя.
Мнение о существовании строгой репродуктивной изоляции между членами комплекса также не находит твердого опытного подтверждения.
Таксономия
Род был первоначально ограниченный по Аполлинарию Семенович Бондарцевым и Рольф Зингер в 1941 году в составе подсемейства Poroideae семьи Polyporaceae. Были включены четыре вида: тип Antrodia calcea , A. crassa , A. xantha и A. lenis . Эти микологи позже независимо друг от друга опубликовали род с латинским описанием (установление действительности публикации в соответствии с правилами номенклатуры ): Зингер в 1944 году и Бондарцев в 1953 году. Исторически сложилось много путаницы в отношении истинной идентичности A. calcea . В настоящее время он классифицируется в роду Antrodia (семейство Fomitopsidaceae ) как Antrodia calceus . Некоторые авторы предпочитают рассматривать Amyloporia , как синоним с антродием . Молекулярное исследование , опубликованное в 2010 году определил , что пять видов , то считаются частью Amyloporia не группа вместе филогенетически , и что Amyloporia не был достоин родового статуса.





Общее название Amyloporia объединяет древнегреческое слово άμυλον («крахмал») и название Poria .