

Описание
Плодовые тела гриба, которые также известны как базидиокарпы, обычно скобки беловатые по краям и темно-коричневые на неровной бугристой верхней поверхности. Однако они также могут принимать ресупинатную форму, состоящую только из белой корочки, которая соответствует нижней стороне скобки. Базидиокарпы достигают 40 см в диаметре и 3,5 см толщины. Плодородная поверхность плодового тела белая, легко покрывается синяками коричневого цвета, с едва заметными порами, по 3-4 на мм. Мякоть, имеющая сильный грибковый запах, в молодом возрасте эластична, но с возрастом становится древесной.
Половые споры, называемые базидиоспорами, образуются в плодородном слое на нижней поверхности базидиокарпов, в то время как конидиоспоры возникают на бесполой стадии и образуются на микроскопических «конидиеносцах», которые прорываются через поверхность дерева-хозяина. Конидиоспоры и базидиоспоры вырабатываются этим грибком, причем последний более важен для заражения хвойных деревьев.
Профилактические меры борьбы и методы лечения насаждений
Для того, чтобы защитить ваши насаждения, необходимо прибегать к использованию профилактических мероприятий. К мерам потенциальной защиты относят:
нужно следить за ними и внимательно оценивать их на предмет новых образований негативного типа;
если же вы увидели, что растение поражено, то стоит его устранить, чтобы недуг не перешел на другие деревья;
насаждения лучше всего делать так, чтобы туда входили как лиственные, так и хвойные деревья
Важно соблюдать разнообразие;
материал для посадки необходимо использовать исключительно качественный, ведь от этого зависит вероятность устойчивости растения к негативным факторам и корневой губки в том числе;
при посадке необходимо добавить в землю такой элемент, как фундазол. Также его следует вводить в почву во время осуществления рубки санитарного типа.
Экология и жизненный цикл
В летнее время выделяются базидиоспоры , первичные инфекционные пропагулы. Эти базидиоспоры переносятся ветровыми течениями на большие расстояния. Они заражают деревья (обычно хвойные) через повреждения, такие как свежескошенные пни. Попав на культю, гриб колонизирует и проникает в корень через мицелий . Heterobasidion annosum перемещается на короткие расстояния от корней инфицированного пня через корневые прививки с другими деревьями. Он также может передаваться через насекомых, питающихся корнями. Поскольку этот грибок не может продвигаться очень далеко через почву, он полагается на корни деревьев, чтобы заразить соседние деревья. В этих корнях он может расти на 0,1–2,0 м в год. Это приводит к распространению грибков и болезней в лесу. Эти болезни образуются, когда деревья умирают и падают, создавая бреши в пологе леса. Эти промежутки влияют на доступную влажность и солнечный свет, изменяя среду обитания растений и животных на лесной подстилке. Spiniger meineckellus , название бесполой стадии этого гриба, образуется на пнях во влажных условиях, и образующиеся конидиоспоры могут жить в почве до десяти месяцев
Роль конидиоспор в инфекционном процессе неизвестна и не считается важной.
Heterobasidion annosum Sensu lato
Heterobasidion annosum Sensu lato представляет собой собрание нескольких видов, вызывающих заболевание корней Heterobasidion и гниль лесных деревьев, а иногда и тех, которые встречаются в ландшафтных насаждениях через Северное полушарие. Эти грибы могут быть сапротрофный или же некротрофный, заселяя неживую сердцевину или живые корни. В качестве хозяев были определены многочисленные виды деревьев, но чаще всего страдают вечнозеленые деревья, такие как сосна. Заболевание корней Heterobasidion и анальная гниль представляют собой серьезную проблему для здоровья леса из-за гибели деревьев, уменьшения объема и качества пиломатериалов, а также уменьшения количества пиломатериалов. связывание углерода. В эту группу видов входят некоторые из наиболее интенсивно изучаемых патогенов лесных деревьев, и они стали первым лесным патогеном базидиомицетов, получившим свою геном секвенирован в 2012.
Heterobasidion annosum Sensu lato включает пять видов, которые различаются предпочтениями хозяина, генетикой, морфология, и географический диапазон. Тесты полового спаривания выявили три группы стерильности: P, S и F, названные в честь их главных хозяев (сосна, ель, и ель ). Пять видов H. abietinum (Тип F), H. annosum sensu stricto (тип P), H. irregulare (P-тип), H. occidentale (S-образный) и H. parviporum (S-образный). Генетический анализ показал, что североамериканские виды, H. irregulare и H. occidentale, отличаются друг от друга и от европейских видов в пределах их межплодовых групп.
МЕТОДИКА
Выделение и идентификация штаммов. Объектами исследования служили 10 штаммов Н1, Н2, Н3, Н4, A-Ha-1, K-Ha-4, Hо-16K,
Hp-8, Т-Ha и 45-2 грибов рода Heterobasidion из коллекции чистых культур лаборатории лесных культур, микологии и фитопатологии
Института леса им. В.Н. Сукачева ФИЦ КНЦ СО РАН (Красноярск, Россия). Штаммы были
выделены в 2014–2018 гг. из плодовых тел и базидиом Heterobasidion, произраставших на различных хвойных породах, либо (в единичном случае) из древесины
поваленного дерева с характерными признаками поражения корневой губкой. Штаммы были
изолированы в чистые культуры на селективных питательных средах: мальт-экстракт-агар
(МЭА), содержащий 0.5% танина и Норкранс-агар (НРКА) . Хранение культур осуществляли на скошенном МЭА при 6°С.
Морфологические признаки изучали у грибов, выращенных на морковном агаре, МЭА и НРКА
при 24 ± 1°С. Микроструктуры оценивали методами фазово-контрастной и светопольной
микроскопии с использованием светового микроскопа Nikon Eclipse Сi с системой фотодокументации
(“Nikon”, Япония) и электронного сканирующего микроскопа Hitachi SEM ТМ-1000 с 10 000
кратным увеличением и разрешением 30 нм (“Hitachi Ltd.”, Япония). Видовую идентификацию
штаммов верифицировали молекулярно-генетическими методами. Секвенирование участков
генетических маркеров ITS (internal transcribed spacer) и TEF-1alpha (transcription elongation factor 1-alpha) проводили на секвенаторе Illumina MySeq (“Illumina”, США) с использованием оборудования
ЦКП “Инновационные технологии защиты растений” Всероссийского научно-исследовательского
института защиты растений (Санкт-Петербург-Пушкин, Россия) и ЦКП “Геномика” (ИХБФМ
СО РАН, Новосибирск, Россия). ДНК выделяли по методу фенол-хлороформной экстракции
[]. Таксономическое положение штаммов и эволюцию определяли с помощью программного
обеспечения MEGA-X . Нуклеотидные последовательности сравнивали с последовательностями известных штаммов
из базы NCBI (Information, U.S. National Library of Medicine).
Условия культивирования грибов, выделение, идентификация и количественное определение
вторичных метаболитов. При изучении продукции вторичных метаболитов штаммы культивировали в глубинных условиях
в колбах (750 мл) со 100 мл среды на шейкере (220 об./мин) в течение 11 сут при 24°С.
При скрининге была использована синтетическая среда НРК []. Для сравнения биосинтетической способности продуктивных штаммов использовали две
органно-минеральные среды. Среда 2 содержала (г/л дистиллированной воды): глюкозу
– 5.0, мальт экстракт – 5.0, KH2PO4 – 0.5, MgSO4 ⋅ 7H2O – 0.5, рН 5.5 []. Среда 3, имела следующий состав (г/л дистиллированной воды): глюкоза – 4.0, дрожжевой
экстракт – 4.0, мальт-экстракт – 10.0, рН 5.5 []. Среды засевали 7-суточным мицелием, выращенным глубинно в среде, содержавшей 3%
мальт-экстракта.
Вторичные метаболиты экстрагировали из фильтрата культуральной жидкости хлороформом
трехкратно при рН 3 и 8. Экстракты анализировали методом ТСХ на пластинках силикагеля
(Silica gel F254, “Merck”, Германия) в системах хлороформ–метанол–25%-ный аммиак (90 : 10 : 0.1) и
хлороформ–ацетон (93 : 7). Метаболиты обнаруживали по поглощению и флуоресценции при
254 и 366 нм. Метаболиты выделяли с помощью препаративной ТСХ (ПТСХ) на пластинках силикагеля (Silica gel F254, “Merck”, Германия) в системах растворителей. Идентификацию выделенных метаболитов
осуществляли на основе сравнения данных УФ-спектроскопии и масс-спектрометрии с литературными
источниками . Подробную информацию о структуре соединений получали при анализе МС/МС спектров
при энергии коллизии 20–40%.
УФ-спектры соединений в метаноле получали на спектрофотометре UV-160А (“Shimadzu”,
Япония). Масс-спектры соединений регистрировали на квадрупольном масс-спектрометре
LCQ Advantage MAX (“Thermo Finnigan”, Германия), используя одноканальный шприцевой
насос для прямого ввода образца в камеру для химической ионизации.
Количественное содержание фоманноксина определяли спектрофотометрически после ПТСХ
на пластинках силикагеля (Silica gel F254, “Merck”, Германия) в метаноле при 280 нм. Расчет проводили, используя калибровочную
кривую для фоманноксина.
Исследование фитопатогенной активности штаммов. Фитопатогенность оценивали по модифицированному методу [] с использованием для культивирования грибов полужидкой питательной среды НРК.
Возбудитель, причины распространения и очаг гриба многолетника
Поражаться данным грибом имеют способность только хвойные породы деревьев. Сюда можно отнести ель, пихту, сосну или же можжевельник. Несомненно, есть и исключения из данного правила, ведь вероятно поражение лиственных деревьев. В таком случае специалисты выделяют березу, дуб или же ясен.
Установлено, что в дикой природе существует несколько так называемых морфологических форм данных грибов. Они в первую очередь все отличаются между собой географическим расположением. Также все имеют разную степень патогенности.
Особенности заболевания лиственницы
Ггриб данного образца редко способен поражать лиственные деревья, но такая вероятность существует. В большей степени это относится к березе, осине и ольхе.
Болезнь ели обыкновенной, пихты и сосны
Гриб корневая губка несет опасность для деревьев и в большей степени это относится исключительно к хвойным породам. Определить подобное поражение можно только по причине наличия на дереве тела гриба. Как правило, такой процесс осуществляется в местности, где влажность повышенная.





![Гетеробазидионсодержание а также heterobasidion annosum sensu lato [ править ]](https://vsegriby.ru/wp-content/uploads/9/7/e/97ed24323c3f2a14f83e05d20796aa53.jfif)

Это многолетняя форма с различной конфигурацией. В общей сложности напоминает просто шляпу, которая одной своею половиной прикреплена к дереву, пню и так далее. Если говорить о поражении данным грибов ели, то он прикрепляется преимущественно к корневой системе.
Что касается поражения соснового дерева, гниль распространяется только по корням и в минимальных случаях может подниматься выше. Когда наступает начальная стадия непосредственного гниения, то корни и их древесина пропитываются так называемой живицей. Она сама по себе становится стекловидной, а также одновременно с этим приобретает красный окрас. Подобная живица имеет способность выходить на поверхность, но со временем это все проходит, а древесина на корне приобретает желтый окрас.
ВАЖНО! Больные деревья серьезно страдают по причине данного гриба. Это обосновывается тем, что у них снижается прирост, появляются побеги укороченного типа, а сама хвоя в тот же момент становится более тусклой и теряет свою привлекательность
На сосняках этот гриб отчетливо выражает усыхание растения.
Если же говорить о поражении данным грибом ели или же пихты, то непосредственная гниль со временем поднимается в комлевую часть ствола. Этот процесс может охватить метр или же два, а в некоторых ситуациях поражает даже все дерево. При таком процессе в корнях начинает свое развитие гниль ядрового и пестрого типа. Фиолетовый окрас древесины свидетельствует о том, что это только первый этап гниения. Со временем цвет меняется и уже может быть красным или же бурым. На дереве можно увидеть белые и продолговатые пятна, что также характерно черными штрихами.
Завершающая стадия подобного гниения сопровождается пустотами в дереве, что напоминает простые ячейки. В таком случае древесина становится полностью дряхлой и окончательно теряет свою прочность. Дерево также истощает крайне негативный запах, который такой же, как и гриб. В процессе разложения гниль может отделиться от здоровой части дерева. Зачастую такой процесс сопровождается исключительно на первом этапе начала гниения корня. Очаги поражения грибом зачастую представляют собой исключительный диффузный или же хаотичный характер. Сперва потрясённые деревья не дают ни малейшего представления, что они уже поражены. Постепенно начинают наблюдаться первые так называемые симптомы. Это проявляется в виде усыхания дерева и значительного сокращения его роста.
ВАЖНО! Корневая губка может поражать как естественные насаждения, так и искусственные. Но в корневых сосняках такое встречается редко
Что же касается ели или же пихты, то там деревья на высоком уровне поражаются как естественного, так и искусственного образца.
Губка корневого типа представляет собой непосредственное заболевание, которое крайне опасное для пород хвойных деревьев. Это обосновывается прежде всего тем, что в случае поражения за быстрый промежуток времени может заразиться большое количество насаждений. В дальнейшем эти деревья массово высыхают, а насаждения другого типа просто теряют свою форму, цвет и привлекательность. Быстрое высыхание говорит и распространении вредителей стволового типа. На территории лесов и насаждений в Российской Федерации данный вредитель встречается весьма часто.
Симптомы и признаки болезни
Симптомы и признаки грибкового заболевания часто обнаруживаются под землей. В H. annosum инфекции вызывают аномальные изменения в структуре корней, доходящие до самого основания дерева. Более половины дерева может погибнуть до того, как человеческий глаз заметит какие-либо симптомы. Базидиокарпия станет видимой в течение полутора или даже трех лет. Эта инфекция вызывает аномальный рост хвои деревьев, бледно-желтую кору, а также увядание и гибель деревьев. Это заболевание корней обычно приводит к тому, что у дерева тонкая крона снизу вверх и изнутри. Деревья рано или поздно умрут. Симптом в ландшафтном масштабе – это кольца мертвых деревьев, находящихся на разных стадиях разложения и гибели, причем самые старые в центре и постепенно молодые движутся наружу. Грибок белой гнили, обнаруженный в корнях, является признаком того, что дерево было поражено H. annosum. Кора меняет цвет по мере прохождения этапов, от бледно-желтого до твердого светло-коричневого, и, наконец, на продвинутой стадии она становится белой с подписями Fomes annosus – посыпанная полоса черных пятен. Другой признак – протекающая часть корня, из-за которой между ним и песком образуется плотная масса.
Симптомы и признаки болезни
Симптомы и признаки грибкового заболевания часто обнаруживаются под землей. В Х. annosum инфекции вызывают аномальное изменение в структуре в корнях , который поднимается до приклада дерева. Более половины дерева может погибнуть до того, как человеческий глаз заметит какие-либо симптомы. Базидиокарпам может понадобиться до полутора или даже трех лет, прежде чем они станут видимыми. Эта инфекция вызывает аномальный рост хвои деревьев, бледно-желтую кору, а также увядание и гибель деревьев. Это корневое заболевание обычно приводит к тому, что у дерева тонкая крона снизу вверх и наизнанку. Деревья рано или поздно умрут. Симптом в ландшафтном масштабе – это кольца мертвых деревьев, находящихся на разных стадиях разложения и гибели, причем самые старые в центре и все более молодые движутся наружу. Грибок белой гнили, обнаруженный в корнях, указывает на то, было ли дерево поражено H. annosum . Кора меняет цвет по мере прохождения стадий: от бледно-желтого до твердого светло-коричневого и, наконец, на поздней стадии она становится белой с характерной чертой Fomes annosus – полосой черных пятен. Еще один признак – протекающая часть корня, из-за которой между ним и песком образуется плотная масса.
Группа порядков афиллофороидные гименомицеты
У представителей этой группы базидии располагаются на поверхности плодовых тел в виде плотного полисадного слоя (гименофора). Плодовые тела крупные, различные по форме — от распростертых паутинистых до плотных консолевидных (иногда с зачатками ножки), однолетние или многолетние. Гименофор может быть гладким, складчатым, бугорчатым, шиповидным, трубчатым, лабиринтообразным. Основные представители этой группы следующие.
Phlebiopsis gigantea (флебия гигантская) характеризуется пленчатым, рыхлым плодовым телом до 50 см длигной. Часто располагается на стволах под корой сухостойных деревьев и пней. Слабый разрушитель древесины (субдеструктор). Гименофор гладкий, мицелий образует ветвистые шнуры. Древесина приобретает бурую окраску.
Telephora terrestris (телефора земляная) вызывает удушье сеянцев в питомниках. Встречается в лесу на подросте. Плодовое тело кожистое, темно-бурое, обволакивает стволик растения, вызывая его ослабление. Использует растение как механическую опору, внутрь тканей растения мицелий не проникает. В питомниках для предотвращения появления и распространения гриба используют приемы агротехники.
Coniophora puteana (пленчатый домовой гриб) разрушает древесину в зданиях и сооружениях. Имеет широкое распространение. Плодовые тела распростертые, мясистые, желто-бурой окраски. Со временем темнеющие. Образует тонкие пленки и шнуры. Гниль бурая, трещиноватая.
Serpula lacrymans (настоящий домовой гриб) — наиболее опасный возбудитель гнили в закрытых помещениях, так как способен использовать влагу, получаемую при разложении древесины. Меры борьбы: профилактика (пропитка древесины антисептиками), замена пораженных деталей и конструкций.
Typhula graminearum — возбудитель выпревания сеянцев (наряду с сумчатым грибом Sclerotinia graminearum). Весной после схода снега на растениях появляются паутинистый, быстро исчезающий мицелий и мелкие темные склероции. Осенью на склероциях образуются булавовидные хрящеватые плодовые тела. Меры борьбы: выполнение требований фитосанитарии и агротехники.
Climacodon septentrionalis (северный ежовик) вызывает белую пластинчатую сердцевинную гниль лиственных пород. Плодовые тела однолетние, в виде плоских шляпок, собранных в черепитчатые группы. Верхняя поверхность шляпок белая, волосистая, позже чернеющая. Гименофор представлен длинными тонкими шипами желтого цвета. Гриб поражает старые деревья (дуб, клен, липу, березу и др.) в садах и парках.




Изоляция
Есть несколько способов изолировать Heterobasidion annosum. Водный агар можно использовать с инфицированной тканью хозяина для получения конидиофоров, которые представляют собой простую или разветвленную часть гиф гриба для устранения гетеробазидиона аннозного. Другой способ изолировать гетеробазидион аннозум – использовать тонкие диски живой заболони из Picea abies. Путем разрезания тонких дисков на чашки Петри, которые используются для культивирования бактерий, и помещения их на влажную фильтровальную бумагу, этот метод позволяет улавливать споры из воздуха, что приводит к бесполой стадии образования грибка на дисках.
Definitioner
- Базидии (Базидия)
Лат. Basidia. Специализированная структура полового размножения у грибов, присущая только базидиомицетам. Базидии представляют собой терминальные (концевые) элементы гиф различной формы и размера, на которых экзогенно (снаружи) развиваются споры.
Базидии разнообразны по строению и способу прикрепления к гифам.
По положению относительно оси гифы, к которым они прикреплены, выделяют три типа базидий:
Апикальные базидии образуются из терминальной клетки гифы и расположены параллельно её оси.
Плевробазидии образуются из боковых отростков и расположены перпендикулярно оси гифы, которая продолжает расти и может образовывать новые отростки с базидиями.
Подобазидии образуются из бокового отростка, повёрнутого перпендикулярно оси гифы, которая после формирования одной базидии прекращает свой рост.
На основе морфологии:
Голобазидии — одноклеточная базидия, не разделенная перегородками (см. рис. А, Г.).
Фрагмобазидии разделены поперечными или вертикальными перегородками, обычно на четыре клетки (см. рис. Б, В).
По типу развития:
Гетеробазидия состоит из двух частей — гипобазидии и развивающейся из нее эпибазидии, с перегородками (см. рис. В, Б) или без них (см. рис. Г).
Гомобазидия не разделяется на гипо- и эпибазидию и во всех случаях считается голобазидией (рис. А).
Базидия — место кариогамии, мейоза и образования базидиоспор. Гомобазидия, как правило, функционально не разделена, и мейоз следует в ней за кариогамией. Однако базидия может быть разделена на пробазидию — место кариогамии и метабазидию — место мейоза. Пробазидия часто является покоящейся спорой, например, у ржавчинных грибов. В таких случаях пробазидия прорастает метабазидией, в которой происходит мейоз и на которой формируются базидиоспоры (см. рис. Д).
См. Кариогамия, Мейоз, Гифа.
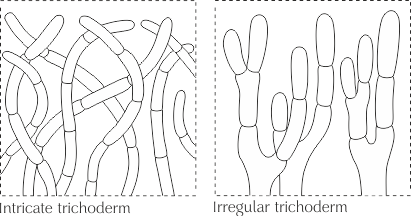
- Триходермы (Триходерма)
Тип кожицы шляпки, состоит из обычно прямых, септированных элементов, расположенных более или менее перпендикулярно поверхности и закладывающихся как на одном, так и на разных уровнях; концы гиф могут быть морфологически видоизменены и представлять собой дерматоцистиды. Поверхность шляпки от бархатистой до почти войлочной.
Лат. Trichoderm.
Триходерма в свою очередь подразделяется на переплетённую триходерму и неправильную триходерму.
Переплетённая триходерма (Intricate trichoderm) – триходерма, состоящая из переплетённых гиф, расположенных непараллельно друг другу и формирующих войлочное опушение.
Неправильная триходерма (Irregular trichoderm) – триходерма, состоящая из нерегулярно ветвящихся гиф.
См. Дерматоцистида, Гифа, Септа.
-
- Апикулюсом (Апикулюс)
1) Короткий вырост на конце споры;
2) Оттянутый короткий отросток базидиоспоры, которым она крепится к стеригме.
См. Гилярный отросток.




Ссылки [ править ]
- ^ а б Корхонен, К. (1978). Группы интерстерильности Heterobasidion annosum. Metsäntutkimuslaitoksen julkaisuja. 94 . ISBN 978-9514003653. OCLC 58493872 .
- ^ a b Отросина, WJ; Гарбелотто, М. (2010). ” Heterobasidion occidentale sp. Nov. И Heterobasidion irregulare nom. Nov.: Расположение североамериканских биологических видов Heterobasidion “. Грибковая биология . 114 (1): 16–25. DOI10.1016 / j.mycres.2009.09.001 . PMID 20965057 .
- ^ а б в Чен, JJ; Корхонен, К .; Li, W .; Дай, YC (2014). «Два новых вида комплекса Heterobasidion insulare на основе морфологии и молекулярных данных». Mycoscience . 55 (4): 289–298. DOI10.1016 / j.myc.2013.11.002 .
- ^ Гарбелотто, М .; Гонтье, П. (2013). «Биология, эпидемиология и борьба с видами Heterobasidion во всем мире». Ежегодный обзор фитопатологии . 51 : 39–59. DOI10.1146 / annurev-phyto-082712-102225 . PMID 23642002 .
- ^ Olson A, Aerts A, Asiegbu F, Belbahri L, Bouzid O, Broberg A, Canbäck B, Coutinho PM, Cullen D, Dalman K, Deflorio G, van Diepen LT, Dunand C, Duplessis S, Durling M, Gonthier P, Гримвуд Дж., Фоссдал К. Г., Ханссон Д., Хенриссат Б., Хиетала А., Химмельстранд К., Хоффмайстер Д., Хёгберг Н., Джеймс Т. Ю., Карлссон М., Колер А., Куэс Ю., Ли Ю. Х., Лин Ю. К., Линд М., Линдквист Е., Ломбард В. , Лукас С., Лунден К., Морин Е., Мюрат С., Парк Дж., Рафаэлло Т., Рузе П., Саламов А., Шмутц Дж., Сольхейм Х., Стольберг Дж., Велез Х., де Фриз Р.П., Вибенга А., Вудворд С., Яковлев И., Гарбелотто М, Мартин Ф, Григорьев И.В., Стенлид Дж (2012). «Понимание компромисса между гниением древесины и паразитизмом из генома грибкового лесного патогена» . Новый фитолог . 194 (4): 1001–13. DOI : 10.1111 / j.1469-8137.2012.04128.x . PMID 22463738 .
- ^ Capretti, P .; Корхонен, К .; Mugnai, L .; Романьоли, К. (1990). «Интерстерильная группа Heterobasidion annosum, специализирующаяся на Abies albaЕвропейский журнал лесопатологии . 20 (4): 231–240. DOI10.1111 / j.1439-0329.1990.tb01134.x .
- ^ Дай, YC; Вайнио, Э.Дж.; Hantula, J .; Niemelae, T .; Корхонен, К. (2002). «Сексуальность и интерстерильность в комплексе Heterobasidion insulareМикологические исследования . 106 (12): 1435–48. DOI10.1017 / S0953756202006950 .
- ^ Дай, YC; Yu, CJ; Ван, ХК (апрель 2007 г.). «Полипоры из восточного Сизанга (Тибет), западного Китая». Annales Botanici Fennici . 44 (2): 135–145. JSTOR 23727602 .
- ^ Дай, YC; Корхонен, К. (2009). Heterobasidion australe , новый полипор, полученный из комплекса Heterobasidion insulareMycoscience . 50 (5): 353–6. DOI10.1007 / S10267-009-0491-3 .
- ^ Токуда, S .; Hattori, T .; Дай, YC; Ota, Y .; Бьюкенен, ПК (2009). «Три вида Heterobasidion (Basidiomycota, Hericiales), H. parviporum , H. orientale sp. Nov. И H. ecrustosum sp. Nov. Из Восточной Азии». Mycoscience . 50 (3): 190–202. DOI10.1007 / S10267-008-0476-7 .
- Перейти ↑ Buchanan, PK (1988). «Новый вид Heterobasidion (Polyporaceae) из Австралазии» . Микотаксон . 32 : 325–337. ISSN 0093-4666 .
| Идентификаторы таксона |
|
|---|
Семейство пукциниевых (Pucciniaceae)
У представителей этого семейства телиоспоры раздельные, иногда соединены в цепочки, с бесцветной ножкой.
Puccinia graminis — возбудитель стеблевой (линейной, черной) ржавчины злаков (II—III—IV) и ржавчины листьев барбариса (0-I). Признаками развития заболевания является образование на листьях злаковых длинных пустул. Пустулы сначала оранжево-желтой окраски, к осени чернеющие (осеннее спороношение). Весенние спороношения — спермогонии и эции оранжевого цвета, чашевидные, развиваются на листьях барбариса. Меры борьбы в питомниках декоративных культур: уничтожение злаков, химическая защита.
Gymnosporangium juniperi — возбудитель ржавчины можжевельника (II-III-IV) и розоцветных пород (0-I). На можжевельнике заболевание вызывает вздутия побегов, на других породах у листьев — красно-оранжевые пятна со столбчатыми эциями. Меры борьбы: удаление посадок от промежуточных хозяев.
Phragmidium mucronatum вызывает ржавчину розы (I-IV), однохозяйный паразит. Эции собраны в группы, красновато-оранжевые. При сильном развитии листья преждевременно опадают, побеги деформируются. Меры борьбы: фитосанитария, химическая защита.
Описание
В первую очередь необходимо выделить второе название корневой губки, которое звучит, как Гетеробазидион многолетний. Что же касается родного латинского языка, то в этом случае Heterobasidion annosum.
Так называемые плодовые тела подобного гриба несимметричны, то есть, он неравномерной формы. В его структуре не предусматривается ножка. Именно по этой причине он крепится к дереву половиной своего основания. Сами по себе плодовые тела могут достигать до 15 см в размерах, а толщина его в максимальной величине составляет 4 см. Если гриб срастается, то может расти в длину до 40 см.
Корневая губка обладает шляпкой в форме раковины. Главная отличительная черта заключается в том, что на своем краю она имеет небольшую выгнутость. Такие грибы очень часто могут срастаться между собой. Также нельзя не вспомнить о неровной поверхности, которой обладают данные шапки. Она покрыта кожицей, которая отличается светло-бурым окрасом.
Края шапки предусматривают контраст от основного окраса, так как имеют оттенок, который приближен к желтому. Если тело гриба молодое, то внутри него преимущественно располагается пробковая мякоть, которая имеет белый цвет или же немного бледный. Такое тело с возрастом начинает темнеть, а также становится более жестким вплоть до деревянного вида. Низ шляпы отображается трубочками блестящего типа, которые слоятся. С каждым годом их становится больше и больше. С самого начала они имеют белый окрас, далее стают более кремовыми, но при надавливании начинают буреть. Способ размножения – белые споры.





