

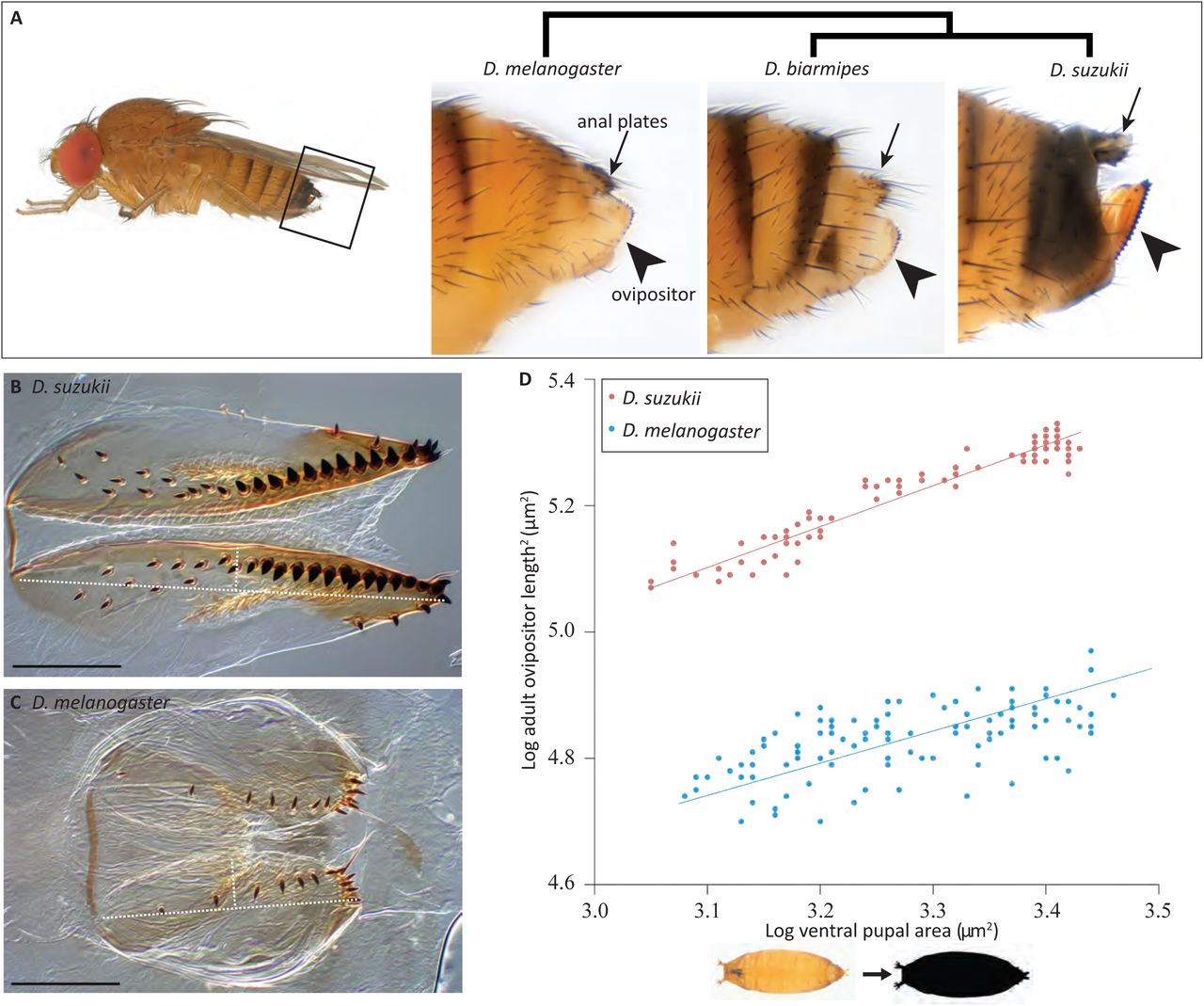
Полёт дрозофилы
Крылья мушки способны колебаться с частотой до 250 раз в секунду. Полёт состоит из прямых поступательных перемещений, чередующихся со стремительными поворотами, называющимися саккадами (saccades). При таких поворотах мушка способна развернуться на 90 градусов менее чем за 50 миллисекунд.
Дрозофилы и, вероятно, многие другие мухи имеют оптические нервы, ведущие напрямую к мускулам крыльев (тогда как у других насекомых они в первую очередь всегда ведут к мозгу), давая возможность очень быстро реагировать.
Долгое время думали, что характеристики полёта дрозофил определяются вязкостью воздуха, а не инертностью их тела. Последние исследования Майкла Дикинсона (Michael Dickinson) и Розалин Сэяман (Rosalyn Sayaman) показали, что когда мушка выполняет поворот, она сначала ускоряется, замедляется на повороте, затем снова ускоряется в конце поворота. Это показывает, что инертность здесь преобладающая сила, также как и в случае с большими летающими животными.
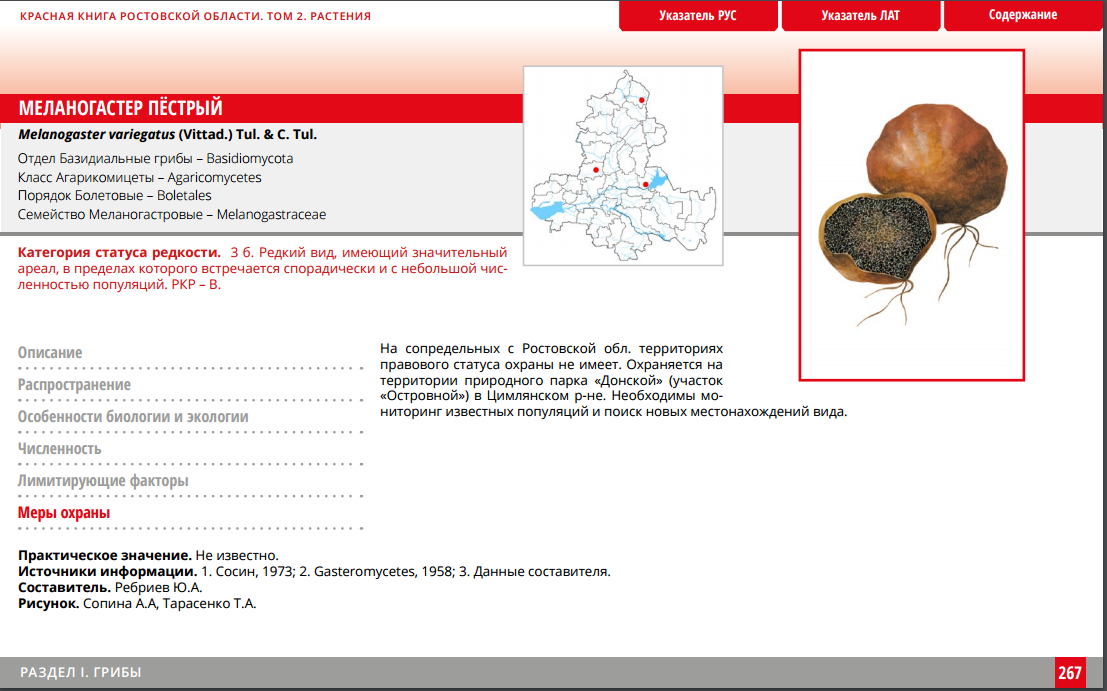
Меланогастер сомнительный (Melanogaster ambiguus)
Синонимы:
- Octaviania ambigua
- Argylium liquaminosum
- Melanogaster klotzschii
Описание
Плодовое тело — гастеромицет, то есть, полностью замкнуто до полного созревания спор. У таких грибов выделяют не шляпку, ножку, гименофор, а гастерокарп (плодовое тело), перидий (внешняя оболочка), глебу (плодоносящая часть).
Гастерокарп 1-3 сантиметра в диаметре, редко до 4 см. Форма от шаровидной до эллипсоидной, может быть правильной либо с неравномерными вздутиями, обычно не разделена на сегменты или лопасти, с нежно-резиновой текстурой в свежем виде. Прикрепляется тонкими, базальными, коричневыми, ветвящимися шнурами мицелия.
Перидий тусклый, бархатистый, сначала серовато-коричневого или корично-коричневого цвета, с возрастом становящийся желтовато-оливковым, с пятнами «синяками» темно-коричневого цвета, в старом возрасте черновато-коричневого цвета, покрытый мелким беловатым налётом. У молодых экземпляров гладкий, затем растрескивается, трещины глубокие, в них видна обнажённая белая трама. В разрезе перидий тёмный, коричневатый.
Глеба изначально белого, беловатого, беловато-желтоватого цвета с голубовато-черными камерами; камеры диаметром до 1,5 мм, более или менее регулярно расположенные, более крупные к центру и основанию, не лабиринтоидные, пустые, желатинизированные со слизистым содержимым. С возрастом, при вызревании спор, глеба темнеет, становится красновато-коричневатой, чёрной с беловатыми прожилками.
Запах: у молодых грибов воспринимается как сладковатый, фруктовый, затем становится неприятным, напоминающим гниющий лук или резину. Англоязычный источник (British truffles. A revision of British hypogeous fungi) сравнивает запах взрослого Меланогастера сомнительного с запахом Scleroderma citrinum (Ложнодождевик обыкновенный), который, по описаниям, напоминает то ли запах сырого картофеля, то ли трюфелей. И, наконец, у созревших экземпляров запах сильный и зловонный.
Вкус: у молодых грибов пряный, приятный
Споровый порошок: чёрный, слизистый.
Под микроскопом
Трамовые пластинки белые, очень редко бледно-желтоватые, тонкие, толщиной 30-100 мкм, плотно сплетенные, гиалиновые, тонкостенные гифы, диаметром 2-8 мкм, не желатинизированные, с зажимными соединениями; межгифальных пространств мало.
Споры 14-20 x 8-10,5 (-12) мкм, изначально яйцевидные и гиалиновые, вскоре переходящие в веретенообразную или ромбовидную форму, обычно с подострой вершиной, полупрозрачные, с утолщенной стенкой от оливкового до темно-коричневого (1-1,3) мкм), гладкие.
Базидии 45-55 х 6-9 мкм, удлиненно-бурые, 2-х или 4 (-6) споровые, часто склеротизованные.
Сезон и распространение
Растёт на почве, на подстилке, под слоем опавших листьев, может быть значительно погружён в почву. Отмечен в лиственных лесах с преобладанием дуба и граба. Плодоносит с мая по октябрь во всей умеренной зоне.
Съедобность
Здесь нет единого мнения. Часть источников указывает Меланогастер сомнительный как однозначно несъедобный вид, часть считает, что гриб можно употреблять в пищу, пока он достаточно молодой (пока глеба, внутренняя часть, не потемнела).
Данных о ядовитости найти не удалось.
Автор данной заметки придерживается принципа «не уверен — не пробуй», поэтому мы осторожно отнесём этот вид к категории несъедобных грибов. Фото: Андрей
Фото: Андрей.
Геном дрозофилы
Геном дрозофилы меланогастер содержит 4 пары хромосом: X/Y пара и три аутосомы, маркируемых как 2, 3 и 4
Четвёртая хромосома точковидная и в ряде исследований её не принимают во внимание; X (или первая), 2 и 3-я хромосомы — метацентрические. Геном состоит из порядка 132 миллионов оснований и приблизительно 13 767 генов
В настоящее время геном секвенирован и аннотирован.
Сходство с человеком
Около 61 % известных человеческих заболеваний имеют узнаваемое соответствие в генетическом коде плодовой мушки, 50 % белковых последовательностей имеют аналоги у млекопитающих. Дрозофилы используются в генетическом моделировании некоторых человеческих заболеваний, включая болезни Паркинсона, Хантингтона и Альцгеймера. Мушка также часто используется для изучения механизмов, лежащих в основе иммунитета, диабета, рака и наркотической зависимости.

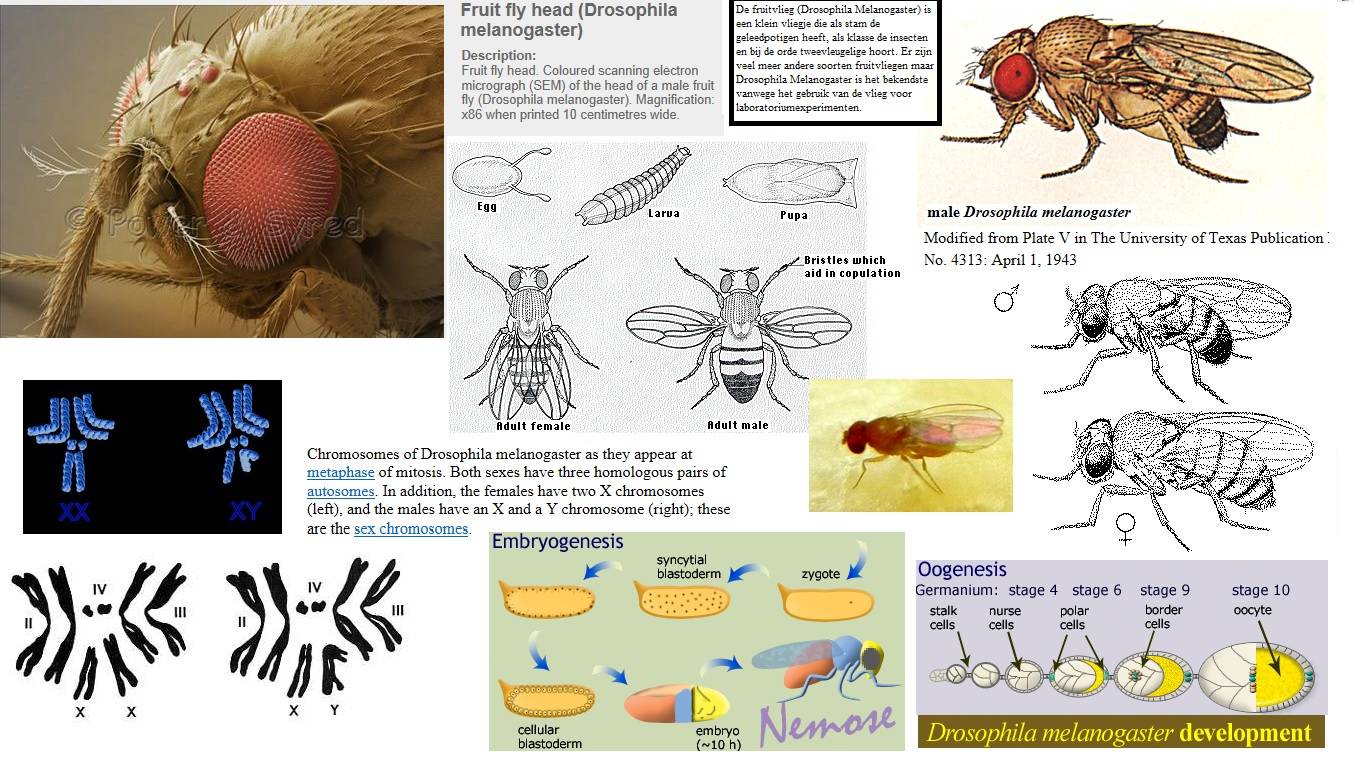







Жизненный цикл
Файл:Drosophila egg.png
Яйцо D. melanogaster
Файл:Drosophila 2nd instar larva.jpg
Личинка D. melanogaster
Жизненный цикл дрозофилы при 25 °C — 10 дней, при 18 °C — примерно в два раза больше. Самки откладывают в загнивающий фрукт или другой органический материал около 400 яиц (эмбрионов), каждое из которых порядка 0,5 мм в длину. Яйца раскрываются через 24 часа. Вылупившиеся личинки растут на протяжении 5 дней, дважды линяя за это время: через 24 и 48 часов после рождения. В течение этого времени они питаются микроорганизмами, разлагающими фрукт, а также и самим сахаром из фрукта. Наконец, личинки закупориваются в пупарий и претерпевают пятидневную стадию метаморфоза, в результате которого возникают взрослые особи.
Первое спаривание у самки происходит не ранее чем через 12 часов после появления из кокона. Самки сохраняют сперму от самцов, с которыми они спариваются, для позднего использования. Именно по этой причине генетики должны отобрать муху женского пола до её первого спаривания (то есть девственную самку) и убедиться, что она спаривается только с конкретным самцом, выбранным для эксперимента. Оплодотворённая самка может быть «возвращена в девственницы» путём длительной инкубации при температуре 10 °C, что убивает сперму, согласно «красной книге» Майкла Эшбёрнера (Michael Ashburner).
Генетика и кариотип
Кариотип – это набор хромосом, который представляет каждая клетка индивидуума после процесса, в котором пары гомологичных хромосом объединяются во время размножения клеток. Этот кариотип характерен для каждого конкретного вида.
Кариотип Drosophila melanogaster Он состоит из одной пары половых хромосом и трех пар аутосомных хромосом. Последние обозначаются последовательно цифрами 2-4. Хромосома 4 намного меньше по размеру, чем остальные ее аналоги.
Несмотря на наличие пары половых хромосом, определение пола у этого вида контролируется отношениями между X-половой хромосомой и аутосомами, а не Y-хромосомой, как это происходит у людей.
Геном, в свою очередь, представляет собой набор генов, содержащихся в этих хромосомах, а у плодовой мушки он представлен примерно 15 000 генов, состоящих из 165 миллионов пар оснований.
Основания азота являются частью ДНК и РНК живых существ. В ДНК они образуют пары из-за конформации двойной спирали этого соединения, то есть основание одной пары спиралей с основанием другой спирали цепи.
Distribution
Latitudinal Distribution Pattern
Latitudinal Range: 23.8° to 22°.
| NorthTemperate | NorthSubtropical | Tropical | SouthSubtropical | SouthTemperate |
Source: AntMaps
Distribution based on Regional Taxon Lists
Oriental Region: India, Myanmar (type locality), Thailand, Vietnam (type locality).Palaearctic Region: China (type locality), Republic of Korea.

Countries Occupied
Number of countries occupied by this species based on AntWiki Regional Taxon Lists. In general, fewer countries occupied indicates a narrower range, while more countries indicates a more widespread species.

Estimated Abundance
Relative abundance based on number of AntMaps records per species (this species within the purple bar). Fewer records (to the left) indicates a less abundant/encountered species while more records (to the right) indicates more abundant/encountered species.

Сомнительный меланогастер
- Гриб редчайший и съедобный, слегка отличается от описанного выше плодового тела. Предпочитает селиться в гордом одиночестве, но иногда встречаются маленькие группки. Искать экземпляры следует, начиная с конца весны, заканчивая серединой осени. Обитание осуществляется в лиственницах с грабами и дубами.
- Гриб клубневидный, как шар, в диаметре может достигать 4 см. Изначально пигментируется коричневатым, позднее становится чёрным или около того. У молодняка поверхность войлочная, у подросших представителей сглаженная. По вкусу приятные, съедобные. Глеба пигментирована чёрно-синим, не распадается в порошковую массу.
- Плодовые тела преимущественно произрастают на почве в лиственных и сосновых лесных массивах. Особенно экземпляры встречаются в грабовых и дубовых. Точно нельзя сказать, но грибники предполагают, что такие плоды образуют микоризу с различными деревьями, под которыми собственно и произрастают.
- Плодоносить плодовые тела начинают с середины весны и вплоть до первых заморозков. Встретить экземпляры можно в европейской части России. Также предполагается, что такой гриб космополит.
- Кроме самой Европы обсуждаемые экземпляры частенько находят в Индии, Северной и Южной Америке, Новой Зеландии. Такой гриб не отличается высокими вкусовыми качествами, но из него можно готовить множество первых и вторых блюд.
Развитие и эмбриогенез
Хромосомное определение пола
Дрозофилам свойственно ХY-определение пола. Важным отличием механизма определения пола дрозофил от человека является то, что на пол влияет не наличие Y-хромосомы, а отношение числа Х-хромосом к числу аутосом. В дальнейшем, имеется в виду гаплоидный набор аутосом (n=4). При отношении равном 1, особь развивается в самку, равном 1/2 – в самца. При нарушениях образуются бесплодные особи: так называемые, интерсексы (если отношение Х-хромосом к аутосомам промежуточное между единицей и 1/2), суперсамцы (отношение меньше 1/2) и суперсамки (отношение больше 1). Наличие Y-хромосомы никак не влияет на пол, но самцы без нее стерильны, так как в ней находятся гены, ответственные за сперматогенез.
| Число Х-хромосом. | Число аутосом в гаплодином наборе. | Отношение числа Х-хромосом к числу аутосом. | Пол (фенотип). |
|---|---|---|---|
| 2 | 2 | 1 | Самка |
| 1 | 2 | 0,5 | Самец |
| 3 | 2 | 1,5 | Суперсамка |
| 2 | 3 | 0,(6) | Интерсекс |
| 1 | 3 | 0,(3) | Суперсамец |
Генетическое определение пола
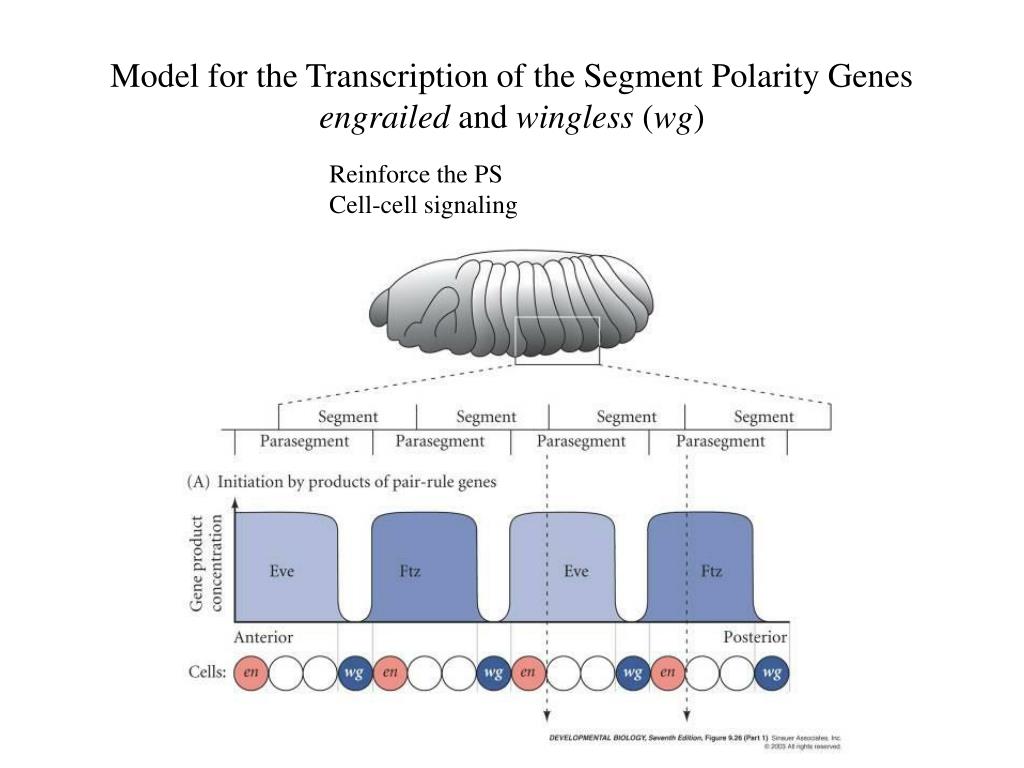
Ключевым геном в определении пола дрозофил является ген sex-lethal (sxl). Именно различие в альтернативном сплайсинге этого гена и порождает различия между двумя полами.
В определении варианта сплайсинга гена sxl участвуют так называемые гены-нумераторы (числителя) (sis-a, sis-b (sisterless-a, sisterless-b), runt (runt)), локализованные в Х-хромосоме, и гены-деноминаторы (знаменателя) (dpn (deadpan) , da (daughterless) , emc (extramacrohaeta) и др.), локализованные в аутосомах. И те, и те являются факторами транскрипции, от соотношения которых зависит алтернативный сплайсинг гена sxl. Активаторы (нумераторы) и репрессоры (деноминаторы) способны образовывать комплексы. В зависимости от отношения количества этих генов, активаторы либо преодолеют действие репрессоров, либо репрессоры подавят активность активаторов. В первом случае (при соотношении X:A=1, генотип женский) ген sxl начнет считываться с проксимального (раннего) промотора. Тогда при альтернативном сплайсинге будет образовываться активный белковый продукт sxl, который является фактором сплайсинга и, связываясь с собственной пре-иРНК, закрепляет такой вариант сплайсинга. В другом же случае, если репрессоры подавили активаторов (отношение X:A=0,5 – генотип мужской), транскрипция sxl начнется с дистального (позднего) промотора При этом образуется неактивный белок, так как не вырезается “мужской экзон” (третий), в котором расположен стоп-кодон UGA, не дающий образоваться полноценной мРНК.
Белковый продукт sxl самки – активный фактор сплайсинга не только для собственной РНК, но и для пре-мРНК гена transformer (tra). У самцов sxl неактивен как фактор сплайсинга, поэтому продукт гена transformer – тоже неактивный белок.
Следующий этап в каскаде – ген Doublesex (dsx). У самок белок tra (совместно с работающим у обоих полов tra-2) модифицирует его сплайсинг, что приводит к образованию белка DsxFem (женский вариант). У самцов такой модификации не происходит, сплайсинг проходит по-другому, и образуется другой белок – DsxM (мужской вариант). Эти белки являются факторами транскрипции, влияющие на активность генов, отвечающих за развитие фенотипического пола.
Дозовая компенсация активности Х-хромосом
У самцов концентрация белков, закодированных в Х-хромосоме, примерно такая же, как и у самок, хотя самих Х-хромосом у них 2 раза меньше. Значит, у дрозофил существуют механизмы, создающие дозовую компенсацию. У человека она тоже существует, но механизмы ее у дрозофилы другие. У человека у самок отключается одна из Х-хромосом, в то время, как у дрозофил повышается в 2 раза интенсивность транскрипции с единственной хромосомы самца. Это видно на цитологических препаратах, где хорошо заметно, что Х-хромосома самца примерно на 25% более рыхлая, чем Х-хромосомы самок. У самцов в ней в полтора раза больше негистоновых белков. Этот эффект вызван активностью определенных белков: msl-1, msl-2, msl-3, mle и гистона H4 с ацетилированным лизином в 16-м положении. Ключевую роль во включении этого механизма играет взаимодействие генов msl-2 и пресловутого sxl. Если белок sxl нормальный (как у самок), он связывает мРНК msl-2 в так называемых UTR-участках, тем самым подавляя её трансляцию. То есть, у самок нет белка msl-2, а у самцов – есть. А при наличии белка msl-2 с ДНК могут связаться и остальные упоминавшиеся белки (msl-1, msl-3, mle и H4Ac16), которые и делают ДНК более рыхлой, что упрощает транскрипцию с Х-хромосомы.



Приготовление и анализ цитологических препаратов
Для получения цитологических препаратов трофоцитов яичников брали 1-1,5 суточных самок дрозофилы, выращенных на стандартном дрожжевом корме. Яичники выделяли в растворе 0,7% NaCl и фиксировали в этанол-уксусном фиксаторе (соотношение 3:1). Окраску производи лактоацетоорсеиновым красителем в течение 10-20 минут. После этого яичники отмывали в 45 % растворе уксусной кислоты и слегка давили на предметном стекле. Препараты хромосом слюнных желез готовили аналогичным способом без предварительной фиксации из личинок третьего возраста. Для анализа пространственного расположения хромосом в ядре у D. santomea готовили недавленые препараты. Для этого окрашенные яичники накрывали покровным стеклом без надавливания для сохранения целостности ядер.
Цитологический анализ трофоцитов яичников проводили при помощи микроскопа Laboval-4 при увеличении 10 х 100 с использованием масляной иммерсии. Анализировали ядра с хорошо развитыми первичными политенными хромосомами. Подсчитывали количество ядер с нарушениями конъюгации гомологичных хромосом (асинапсисами).
У межлинейных гибридов Fl Canton S х Berlin, Oregon R х 7 J А х W1118 ядра, имеющие асинапсисы, относили к одному классу. При анализе трофоцитов гетерозигот по хромосомным перестройкам выделяли три класса ядер: 1) ядра без видимого нарушения спаривания гомологов; 2) ядра с небольшими участками асинаптирования; 3) ядра со значительными нарушениями конъюгации гомологичных хромосом.
При оценке контрольной линии yellow и гетерозигот по транслокации Т(1;2)1Е; 23C-D ко второму классу относили ядра с асинапсисом одного из пяти плеч. При анализе гетерозигот по парацентрическим инверсиям в Х-хромосоме: 1) In(l)7B-C; 12E-F; 2) 1п(1)1Е;20В-С\ и 3) Іп(1)4В-С; 20С во второй класс включали ядра с нарушением конъюгации гомологов в 1-2 плечах, гетерозигот по перицентрическим инверсиям (In(2LR) Су О; In(2LR) Pm) — в 1-3 плечах. Это связано с тем, что наличие в генотипе инверсии в гетерозиготном состоянии, как правило, нарушает спаривание гомологов. Таким образом, к третьему классу относили ядра, в которых имелось более одного асинапсиса, не связанного с наличием в плече инверсии. Достоверность различий между опытами и контролем по количеству ядер с асинапсисами рассчитывали с помощью критерия Хи-квадрат. Количество проанализированного материала представлено в таблице 2.3.1.
Для получения хромосомных перестроек в Х-хромосоме на основе линии yellow брали самцов и облучали гамма-лучами в дозе 3000 Gr (установка «Игур-1»). Затем их скрещивали с виргинными самками линии, несущей цепочку маркеров в Х-хромосоме – у ее cv ct vf. Полученных от скрещивания дочерей, имеющих одну облученную Х-хромосому, а вторую — с цепочкой маркеров, сажали индивидуально с самцами у ее cv ct v f Отбирали самок, в потомстве которых не было кроссоверных сыновей, или их было мало. Их сыновей, которые не несли цепочку маркеров, вводили в культуру, скрещивая с самками линии Muller-5 и линии, несущей сцепленные Х-хромосомы. Полученные культуры проверяли на наличие перестроек цитологически.
Полученные хромосомные перестройки поддерживали в культуре путем скрещивания самцов yellow с перестройкой в Х-хромосоме с самками со сцепленными Х-хромосомами C(1)DX, у2 wf. Чтобы исключить влияние аутосом в тестах по действию перестроек на проявление леталей, самок исходной линии C(1)DX, у2 w/в течение десяти поколений скрещивали с самцами линии yellow. Такая процедура обеспечивает замену аутосом исходной линии C(1)DX, у2 w f на аутосомы линии yellow. Далее самцов с перестройкой скрещивали с этими самками. Таким образом, самцы с хромосомными перестройками отличались от фондовых самцов yellow только наличием перестройки.
Для получения самок-гетерозигот по инверсиям и транслокации самок фондовой линии yellow скрещивали с самцами, несущими перестройки. Виргинных самок F1 использовали для тестирования мутаций.
Ссылки
- М. Эшбёрнер и T.R.F. Райт (1978). Генетика и биология дрозофилы. Том 2а. Академическая пресса.
- М. Эшбернер, К.Г. Голич и Р. Хоули (2005). Дрозофила: Лабораторный справочник 2-е издание. Лабораторный пресс Колд-Спринг-Харбор.
- Дрозофиламеланогастр. В Википедии. Восстановлено с en.wikipedia.org.
- Х. Гонсалес (2002). Сравнительная эволюция хромосомных элементов у пола Дрозофила. Докторская диссертация. Автономный университет Барселоны, Испания.
- М. Швентнер, Д.Дж. Combosch, J.P. Нельсон и Дж. Гирибет (2017). Филогеномное решение происхождения насекомых путем выяснения отношений ракообразных и гексапод. Текущая биология.
- С. Ямамото, М. Джайсвал, В.-Л. Чанг, Т. Гамбин, Э. Караджа… и Х.Дж. Беллен (2015). К Дрозофила генетический ресурс мутантов для изучения механизмов, лежащих в основе генетических заболеваний человека. Ячейка
Ссылки
- М. Эшбернер и TRF Райт (1978). Генетика и биология дрозофилы. Том 2а. Академическая пресса.
- М. Эшбернер, К.Г. Голич и Р.С. Хоули (2005). Дрозофила: лабораторный справочник 2-е издание. Лабораторный пресс Колд-Спринг-Харбор.
- Drosophila melanogaster. В Википедии. Восстановлено с en.wikipedia.org.
- Х. Гонсалес (2002). Сравнительная эволюция хромосомных элементов в роде Drosophila. Докторская диссертация. Автономный университет Барселоны, Испания.
- М. Швентнер, DJ Combosch, JP Nelson & G. Giribet (2017). Филогеномное решение происхождения насекомых путем выяснения отношений ракообразных и гексапод. Текущая биология.
- С. Ямамото, М. Джайсвал, В.-Л. Чанг, Т. Гамбин, Э. Караджа… и Х. Дж. Беллен (2015). Генетический ресурс мутантов дрозофилы для изучения механизмов, лежащих в основе генетических заболеваний человека. клетка
История
Первые исследователи, нанявшие Drosophila melanogaster Объектом генетических исследований был Томас Хант Морган и его сотрудники в 1910 году. Эти исследователи из Колумбийского университета (США) изучали насекомых в лаборатории, известной как «комната мух».
Питательной средой, которую Морган и его сотрудники использовали для содержания мух, были молочные бутылки. Сегодня используются более изощренные средства, в том числе мацерированные фрукты и химические консерванты для их поддержания.
Короткий жизненный цикл и большое количество потомства, которое может быть получено за короткое время, позволили использовать эту муху для понимания механизмов, связанных с наследованием, связанным с полом, фенотипической экспрессией за счет множественных аллелей, а также взаимодействием между генами. а также для разработки генетических карт.
Благодаря важности для генетических исследований, это был один из первых организмов, геном которого был изучен. В 2000 году стало известно, что Drosophila melanogaster благодаря усилиям государственных и частных институтов он обладал более чем 13 500 генами
Спустя более века после первых исследований Моргана и его коллег, плодовая муха по-прежнему широко используется в качестве генетической модели для понимания различных заболеваний человека, от болезней обмена веществ и иммунной системы до нейродегенеративных заболеваний, таких как болезнь Паркинсона и Альцгеймера.
характеристики
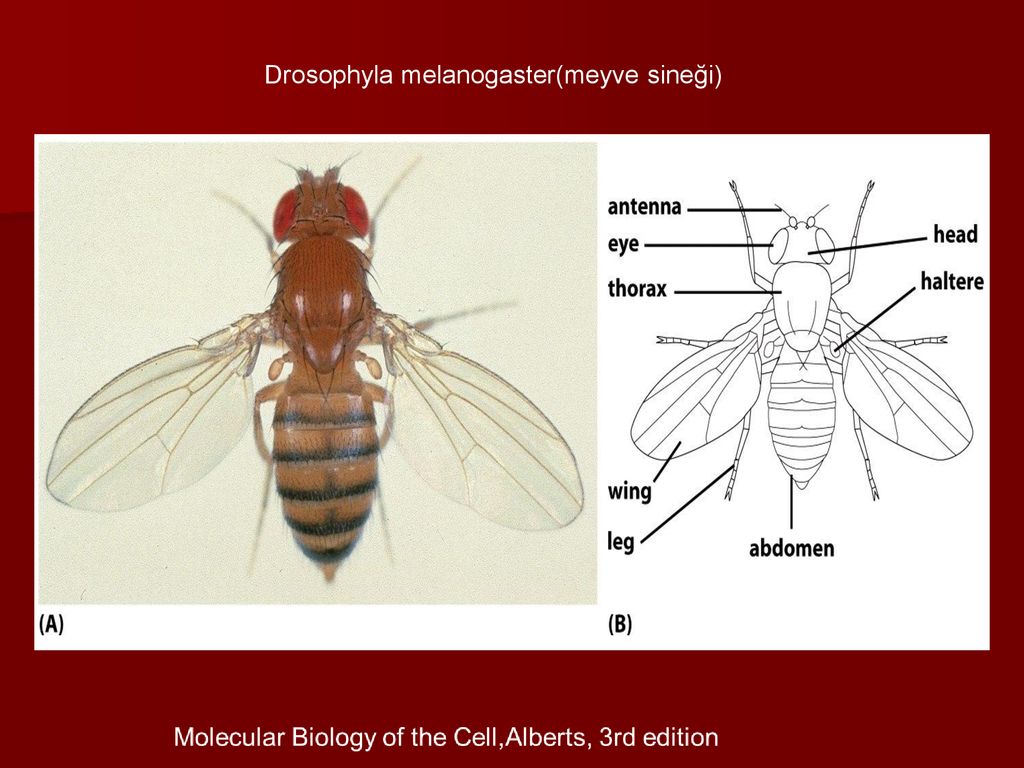
Drosophila melanogaster Это двукрылое насекомое, то есть у него одна пара перепончатых крыльев вместо двух пар, как у других насекомых. Это раздельнополый организм, то есть он представлен раздельными полами. Кроме того, это половой диморфизм.
Размер этого вида составляет около 3 мм, самка немного крупнее самца. Его тело состоит из трех тагмат (областей): головы, груди и живота. Количество сегментов головы (6) и грудной клетки (3) одинаково у самцов и самок, тогда как количество сегментов брюшка у самок (7) больше, чем у самцов (6).
Шесть головных сегментов сливаются, и первый распознается, потому что он несет усики, которые состоят из трех частей, называемых артефактами. Три сегмента грудной клетки также слиты, и на каждом из них есть пара ног. Крылья прикреплены ко второму сегменту грудной клетки.
Различие в количестве сомитов брюшной полости у обоих полов происходит из-за слияния двух последних сегментов у самцов.
Как видно из названия, у мух этого вида брюшко темного цвета, однако существуют мутации, которые могут повлиять на количество и распределение пигментов в их теле, придавая им желтый или полностью черный цвет.
Характерной чертой этого вида на хромосомном уровне является то, что они представляют гигантские (полигенные) хромосомы в слюнных железах. Полигенные хромосомы – это хромосомы, которые подверглись 10 или более сериям репликаций ДНК, но остаются в состоянии интерфейса, то есть деления клеток не происходит.
Таксономия и классификация
Плодовая муха – насекомое. Традиционное расположение членистоногих включает насекомых (или гексапод) вместе с многоножками, многоножками, симфиланами, пауроподами и ракообразными в составе группы челюстных членистоногих.
Более поздние классификации исключают ракообразных из группы и помещают остальных в подтип Uniramia. Однако молекулярные исследования показывают, что насекомые связаны с некоторыми низшими ракообразными, последние являются полифилетической группой.
В любом случае плодовые мухи принадлежат к отряду Diptera, подотряду Brachycera и к семейству Drosophilidae. Пол Дрозофила Он состоит из около 15 подродов и около 2000 видов.
Виды D. melanogaster был описан Майгеном в 1830 г. и относится к подроду Софофора, который насчитывает около 150 видов, разделенных на 10 различных подгрупп, принадлежащих к D. melanogaster в подгруппу меланогастр
Жизненный цикл
Жизненный цикл Drosophila melanogaster Он короткий, в среднем, длится от 15 до 21 дня. Однако его долговечность может варьироваться в зависимости от условий окружающей среды, в основном из-за температуры окружающей среды, в которой он находится.


Например, мухи, выращенные в среде с относительной влажностью 60%, имеют жизненный цикл около 10 дней, если они содержатся при температуре 25 ºC; а при температуре всего 20 ºC – 15 дней. Однако при 29 ° C они могут жить 30 дней, если условия влажности достаточны.
После спаривания самка откладывает до 500 яиц длиной примерно 0,5 мм, из которых вылупляются личинки через 24 часа эмбрионального развития. Развитие личинок состоит из трех стадий продолжительностью около суток каждая.
После личиночной стадии продолжается стадия куколки, которая длится 4 дня. Куколка претерпевает полную метаморфозу и уступает место взрослой особи, которая достигает половой зрелости через 12 часов после выхода из куколки.
Мутации в крыльях
Развитие крыльев у Drosophila melanogaster кодируется хромосомой 2. Мутации в этой хромосоме могут вызывать аномальное развитие крыльев либо по размеру (рудиментарные крылья), либо по форме (изогнутые или изогнутые крылья).
Первая из этих мутаций является рецессивной, то есть для того, чтобы проявить себя фенотипически, мутантный ген должен быть унаследован от отца и матери одновременно. Напротив, мутантный ген изогнутых крыльев является доминантным, однако он проявляется только тогда, когда носитель является гетерозиготным, поскольку гомозиготы нежизнеспособны.
Возможно также появление полностью бескрылых организмов.
Генетика и кариотип
Кариотип – это набор хромосом, который представляет каждая клетка отдельного человека после процесса, в котором пары гомологичных хромосом соединяются во время размножения клеток. Этот кариотип характерен для каждого конкретного вида.
Кариотип Drosophila melanogaster состоит из одной пары половых хромосом и трех пар аутосомных хромосом. Последние обозначаются последовательно цифрами 2-4. Хромосома 4 намного меньше по размеру, чем остальные ее аналоги.
Несмотря на наличие пары половых хромосом, определение пола у этого вида контролируется отношениями между X-половой хромосомой и аутосомами, а не Y-хромосомой, как это происходит у людей.
Геном, в свою очередь, представляет собой набор генов, содержащихся в этих хромосомах, а у плодовой мушки он представлен примерно 15 000 генами, состоящими из 165 миллионов пар оснований.
Основания азота являются частью ДНК и РНК живых существ. В ДНК они образуют пары из-за конформации двойной спирали этого соединения, то есть основание одной спирали попадает в пару с основанием другой спирали цепи.
Мутации в крыльях
Развитие крыльев в Drosophila melanogaster он кодируется хромосомой 2. Мутации в этой хромосоме могут вызывать аномальное развитие крыльев либо по размеру (рудиментарные крылья), либо по форме (изогнутые или изогнутые крылья).
Первая из этих мутаций является рецессивной, то есть для фенотипического проявления мутантный ген должен быть унаследован от отца и матери одновременно. Напротив, мутантный ген изогнутых крыльев является доминантным, однако он проявляется только тогда, когда носитель является гетерозиготным, поскольку гомозиготы нежизнеспособны.
Возможно также появление полностью бескрылых организмов.





