

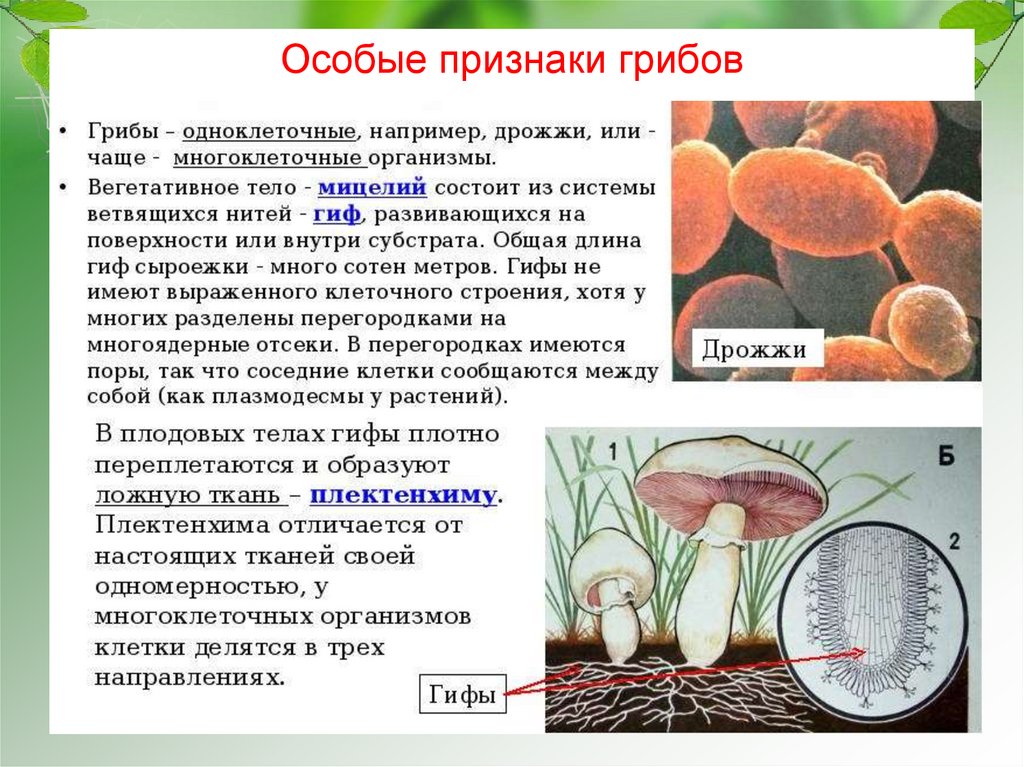
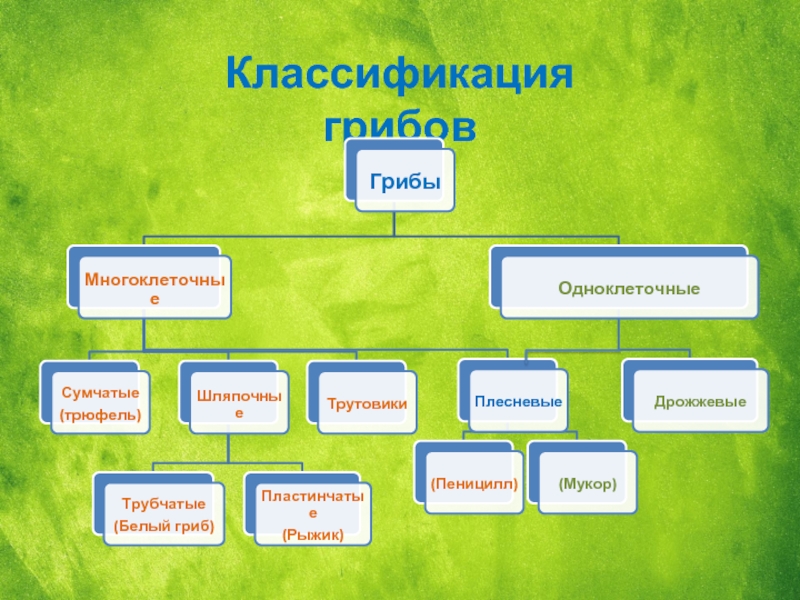
Морфология
Плодовые тела – отсутствуют.
Устоспоры – толстостенные споры, из которых образуются базидии. Устоспоры возникают из ранее существовавшего мицелия по типу хламидиоспор.
Мицелий – дикариотический, распространяется по межклеточным промежуткам тканей растений-хозяев, в клетки проникает при помощи гаусторий. .
Биология
Представители класса Устилягономицеты (Ustilaginomycetes) – паразитные микроскопические грибы. Они поражают цветки (тычинки, завязи) семена, вызывая их разрушение. Кроме того, могут поражаться листья, стебли, корни. На них появляются характерные вздутия в виде полос или галлов темного цвета. Может наблюдаться деформация всего растения или отдельных органов.
Половой процесс – соматогамия. Он протекает в виде копуляции базидиоспор, продуктов их почкования или в виде копуляции гаплоидных клеток базидий (без образования базидиоспор). В результате формируется дикариотичный мицелий.
Такая форма полового процесса обусловлена паразитическим образом жизни головневых грибов и максимальной редукцией гаплоидных стадий, как нежизнеспособных в этих условиях. Характерно, что полученные на стерильных искусственных средах головневые грибы существуют в гаплоидном состоянии.
Биология
Представители класса Зигомицеты (Zygomycetes) почти исключительно приспособлены к сухопутному сапротрофному образу жизни. Виды класса вызывают порчу продуктов и различных материалов. Многие широко распространены в почве, участвуют в почвообразовательных процессах, накоплении гумуса.
В цикле развития отсутствуют подвижные стадии. Основная часть жизненного цикла приходится на гаплоидную фазу. Может наблюдаться гетероталлизм и гомоталлизм.
Вегетатативное размножение
Вегетативное размножение осуществляется частями вегетативного тела таллома или при помощи видоизменений мицелия: столонов, хламидоспор.
Бесполое размножение
Бесполое размножение осуществляется неподвижными эндогенными спорангиоспорами, образующимися в спорангиях или, гораздо реже, экзогенными конидиями. В ряде семейств прослеживается эволюция бесполого размножения – видоизменение специализированных структур, анаморф, от эндогенных спорангиоспор через малоспоровые спорангии к экзогенным конидиям. Это, скорее всего, связано с переходом зигомицетов к наземному образу жизни.
Некоторые виды характеризуются образованием структур промежуточного типа между спорангиями и конидиями – спорангиол. Это мелкие спорангии без колонки. В спорангиенолах количество спор не превышает десяти. При неблагоприятных условиях их число сокращается до одной. Спорангиенолы сходны с конидиями. Есть виды, у которых спорагиеспоры отсутствуют, их заменяют конидии.
Половой процесс
Половой процесс – зигогамия. Он известен не у всех видов класса. Это слияние недифференцированного на гаметы содержимого двух клеток, отделяющихся перегородками от несущих гиф. Сливаются одноядерные и многоядерные клетки. На месте слияния формируется зигота или покоящаяся зигоспора, одетая толстой, скульптурной оболочкой. На зиготе остаются участки гиф, отделяющих гаметангии. Они морфологически отличаются от остального мицелия и называются суспензорами или подвесками, и часто приподнимают зиготу над субстратом.
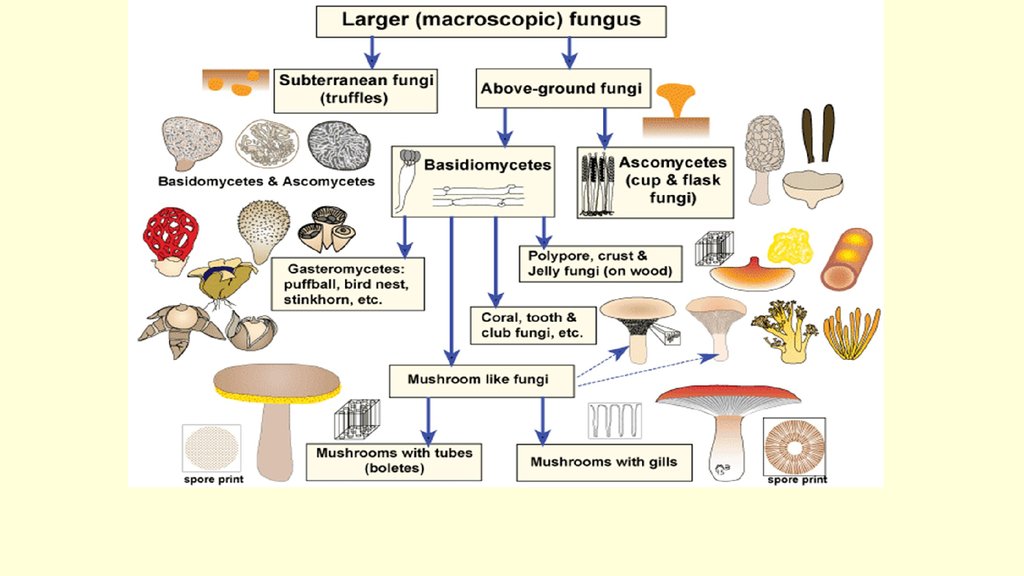
Перидий аскомикот
Образование перидия у аскомикот (сумчатых грибов) характерно для видов с настоящими аскокарпами (плодовыми телами). Перидий в данном случае образуется только при плазмогамии. При этом гаплоидные вегетативные гифы оплетают аскогенные гифы и аски, образуя плотную покровную ткань.
Перидий выделяют у клейстотециев и перитециев. Апотеции обычно не имеют перидия. Исключение – виды порядка Трюфелевые (Tuberales), образующие вторично замкнутые в зрелом состоянии плодовые тела.
Клейстотеции у большинства видов Мучнисторосяных грибов (порядок Erysiphales) видны невооруженным глазом, как мелкие точки, вначале белого цвета, затем желтеющие и в конце темно-коричневые или черные. Окраска связана со степенью зрелости перидия.
От наружных клеток перидия клейстотеций отходят придатки (отростки, аппендиксы) различного строения, представляющие собой мертвые клетки с сильно утолщенными стенками.
Немногие примитивные представители класса Эуаскомицеты (Euascomycetidae) образуют аски группами или пучками на мицелии. Такие аски не окружены перидием либо перидий плодового тела имеется в форме рыхлого, просвечивающегося сплетения гиф. К таким видам относятся представители родов Биссохламис (Byssochlamys) и Аскодесмис (Ascodesmis).
Перитеции характеризуются хорошо развитым, плотным перидием. Плодовые тела типа «перитеции» характерны для класса Сордариомицеты (Sordariomycetes).
Морфология
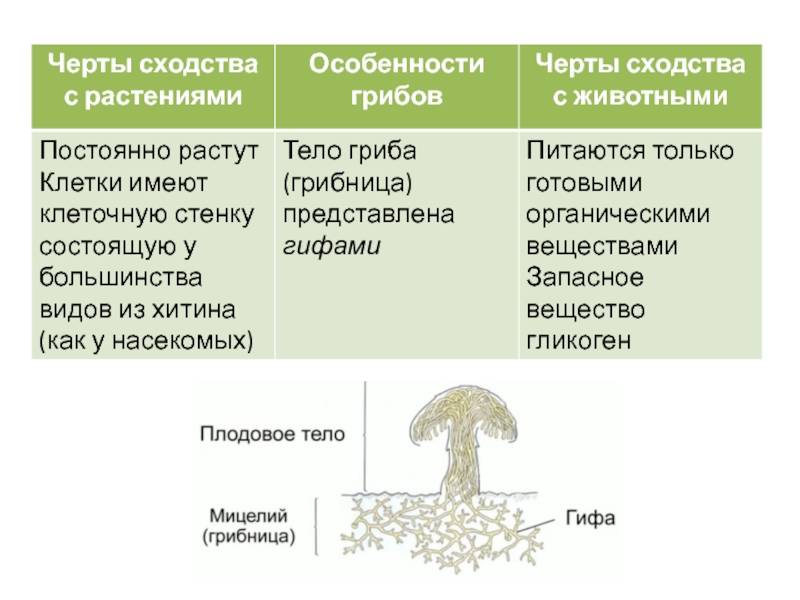
Вегетативное тело – обильно разветвленный многоядерный неклеточный мицелий, субстратный и воздушный. У некоторых видов в зрелом состоянии формируются клеточные перегородки, разделяющие мицелий на отдельные многоядерные фрагменты.



Подвижные стадии – отсутствуют.
Ризоиды, столоны – поддерживающие и проводящие структуры мицелия. Морфология зависит от видовой принадлежности.
Гаусторий, аппрессорий – инфекционные структуры мицелия. Морфология зависит от видовой принадлежности.
Хламидоспоры – органы вегетативного размножения. Морфология зависит от видовой принадлежности.
Спорангиеспоры, спорангиолии, конидии – органы бесполого размножения. Внешний вид зависит от видовой принадлежности.
Зигота (покоящаяся зигоспора) – структура полового размножения, одетая в толстую скульптурную оболочку с суспензорами или подвесками.
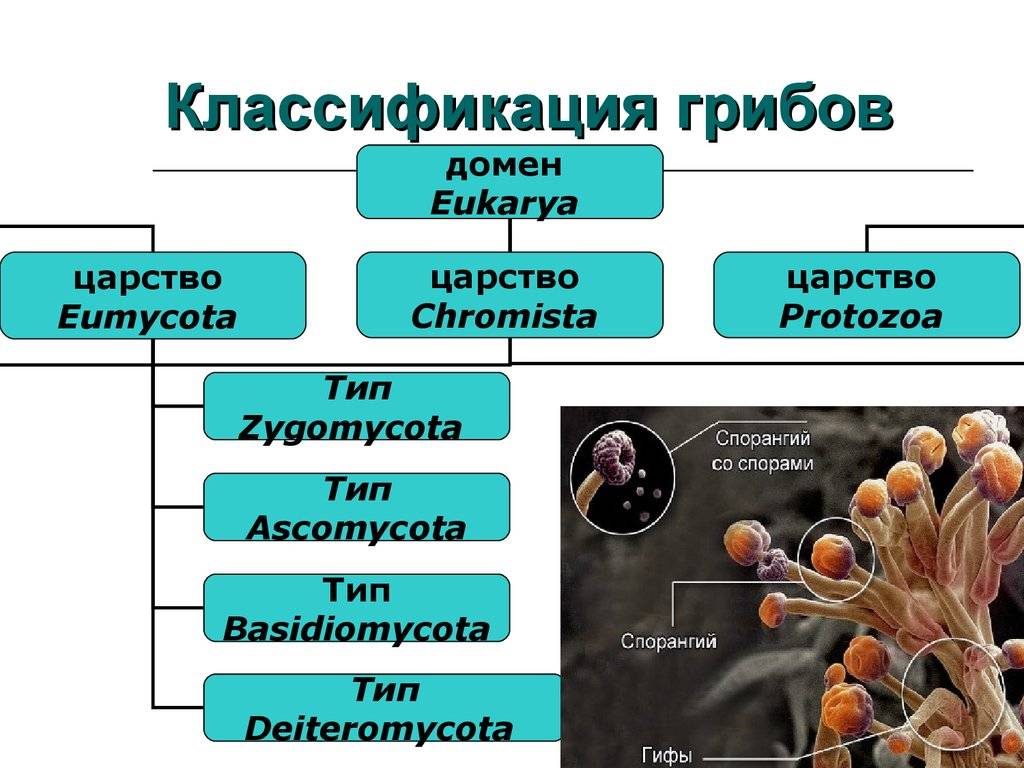
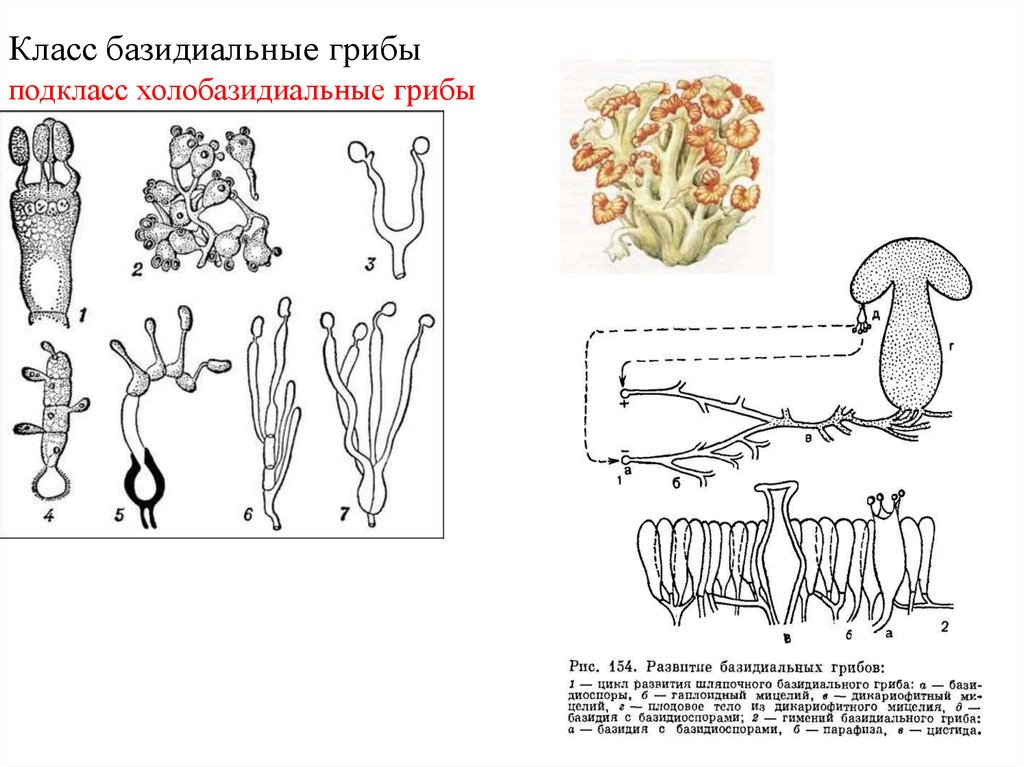
Место класса Устилягономицеты в макросистеме грибов
На современном этапе развития систематики грибов и грибоподобных организмов тип (отдел) Базидиомикота (Basidiomycota) разделен на 3 подотдела:
- Подотдел Pucciniomycotina (Пукциниомицеты).
- Подотдел Ustilaginomycota (Устилагиномицеты).
- Подотдел Agaricomycotina (Агарикомицеты).
Подотдел Ustilaginomycota (Устилагиномицеты) включает два класса:
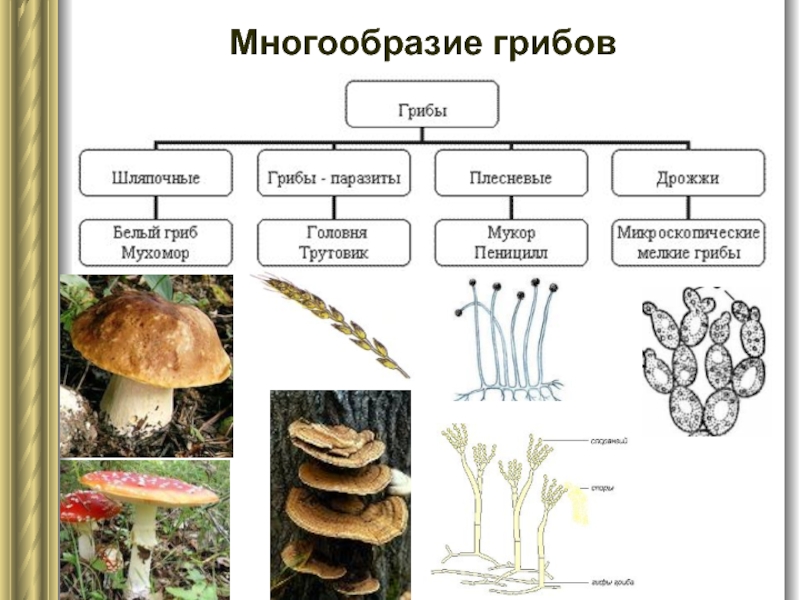
- Класс Ustilaginomycetes (синоним Ustomycetes – Устомицеты) – головневые грибы.
- Класс Exobasidiomycetes (Экзобазидиомицеты).
На современном этапе класс Устилягономицеты (Ustilaginomycetes) включает два порядка:
- Порядок Urocystidales.
- Порядок Головневые (Ustilaginales).
Кроме того, семейство Тиллециевые (Tilletiaceae), расположенное ранее в пределах порядка Головневые (Ustilaginales) (Головневые), в настоящее время предложено поднять до ранга порядка Тиллециевые (Tilletiales) и отнести к классу Экзобазидиомицеты (Exobasidiomycetes).
В литературе встречается вариант систематики, при котором класс Головневые грибы делят на три порядка: Устилягинальные (Ustilaginales), Тиллециальные (Tilletiales), Экзобазидиальные (Exobasidiales) .
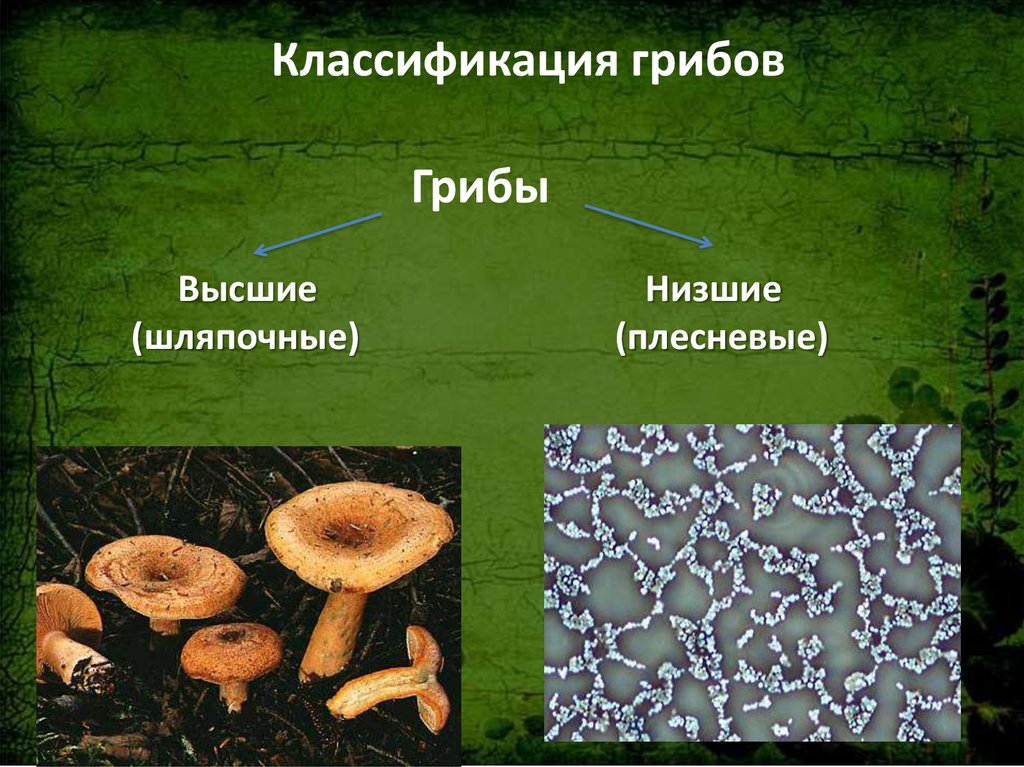
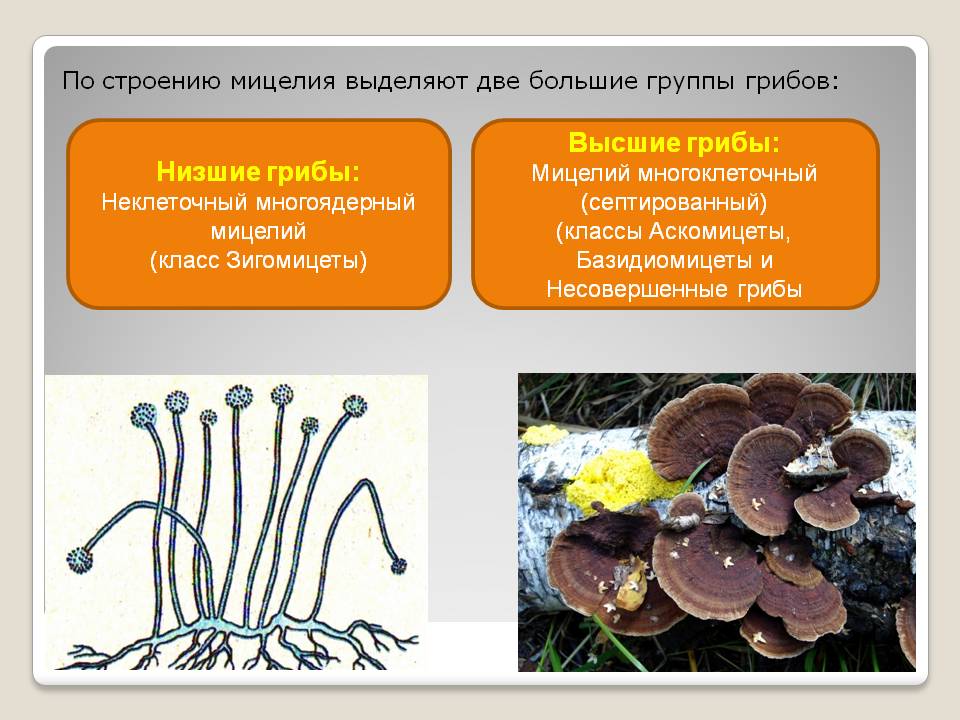
Класс зигомицеты (низшие грибы)
Зигомицеты представлены быстрорастущим видами, обычно обитающими в почве.
При культивировании in vitro образуют обильный сероватый или белый воздушный мицелий.
Их гифы не имеют перегородок либо септированы части1 но. Размножаются половым и бесполым путём.
Бесполое размножение реализуется через образование спорангиофоров со спорангиями.
Половое размножение приводит к образованию зигот — зигоспор. Поражения человека носят выраженный оппортунистический характер.
Их возбудителями могут быть представители родов Absidia, Mortierella, Mucor, Rhizopus, Entomophthora, Conidiobolus и Basidiobolus.
Для бесполого размножения образуют спорангии или спорангиоли (многочисленные спорангии с 2-3 спорами).
Многие распространяются столонами — выбрасываемыми в воздушную среду длинными гифами,
которые, находя подходящий субстрат, выпускают ризоиды и дают начало новой колонии.
Мицелий зигомицетов имеет два знака.
При контакте мицелия разных знаков, формируется зигота из которой прорастает спорангий с гаплоидными спорами.
Зоопаги
Порядок Zoopagales включает в себя хищные грибы , обладающие клейкими гифамами и ловчими кольцами
и питающиеся простейшими, нематодами и мелкими личинками насекомых.
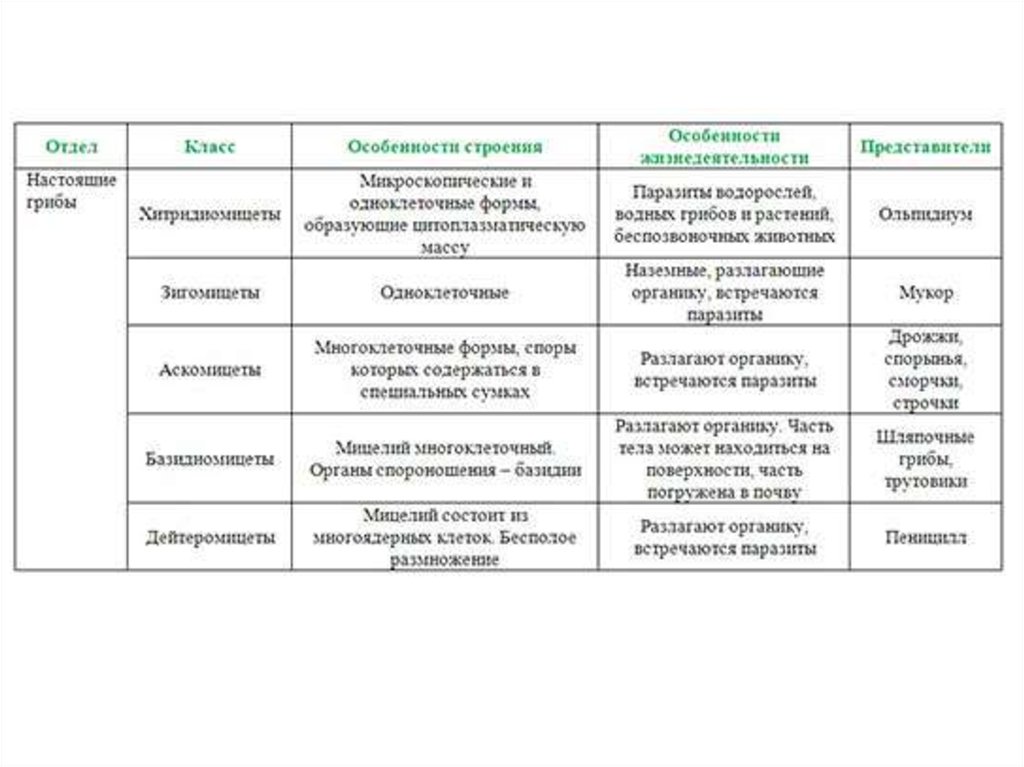
Класс дейтеромицеты (несовершенные грибы)
Дейтеромицеты (от греч. deuteros, вторичный, + mykes, гриб) не образуют настоящую филогенетическую группу,
а выступают в качестве таксономической «свалки», куда помещают виды, у которых половая (совершенная) стадия
размножения отсутствует либо не выявлена.
Их классификация основана на формах спороношения или других внешних признаках и служит только практическим целям.
Для них установленным считают лишь бесполое размножение, поэтому дейтеромицеты также известны
как несовершенные грибы (Fungi imperfecti).
По морфологическим признакам большинство дейтеромицетов сходно с аскомицетами.
Большая часть возбудителей микозов человека входит в группу несовершенных грибов.
Тело дейтеромицет состоит из расчлененных прозрачных или окрашенных многоклеточных гиф и иногда из почкующихся клеток.
Размножаются исключительно бесполым путем, при котором образование конидий происходит
на изолированных или расположенных группами конидиеносцах или специальных образованиях, называемых пикнидами.
К этому классу относятся три порядка: Sphaeropsidales, Melanconiales и Hyphomycetales (Moniliales),
представители которых широко распространены в почве.
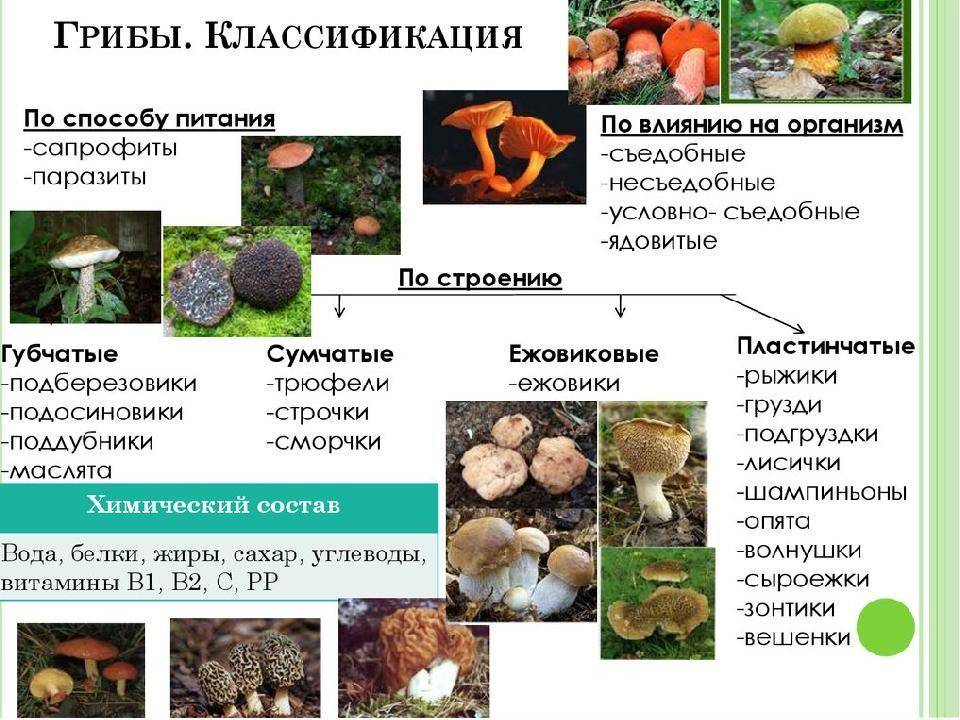
Порядок сферопсидные (Sphaeropsidales)
Грибы порядка Sphaeropsidales характеризуются конидиями, которые образуются в пикнидах,
остающихся закрытыми или открывающихся наружу порами или трещинами.
Сюда входит род Phoma и др. Виды рода Phoma образуют микоризу с корнями некоторых растений.





Порядок Melanconiales
В порядок Melanconiales входят организмы, которые не имеют пикнид.
Конидии расположены на конидиеносцах, соединенных в особые образования – ацервулы.
Порядок гифомицетальные (Hyphomycetales, или Moniliales)
Грибы порядка Hyphomycetales имеют расчлененные, разветвленные, прозрачные или темно окрашенные гифы.
Их весьма разнообразные конидии находятся на конидиеносцах, последние расположены по одному или группами.
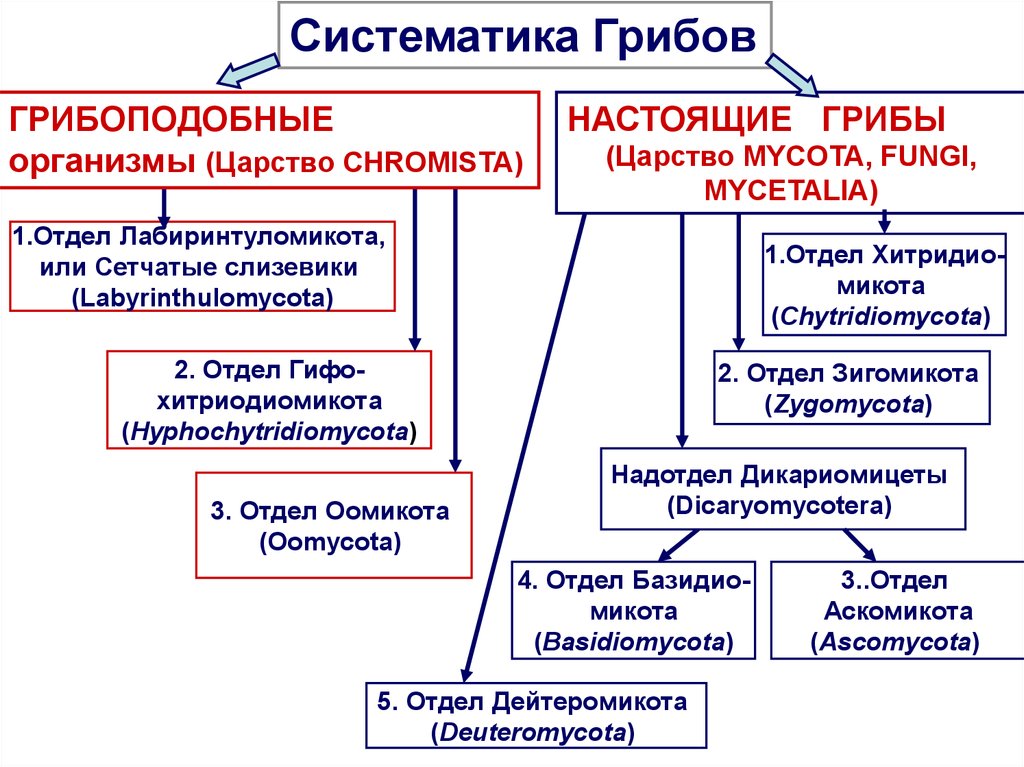
Место класса Зигомицеты в макросистеме грибов
Тип (отдел) Зигомикота (Zygomycota) – один из семи типов (отделов) царства настоящих грибов Fungi, Mycota. Кроме класса Зигомицеты (Zygomycetes) к типу Зигомикота относится дискуссионный класс Трихомицеты (Trichomycetes). В него входят обитатели кишечника или хитиновых покровов насекомых, эндосимбионты или сапротрофы (ассоцианты).
Традиционно класс Зигомицеты делится на несколько порядков (от тех до восьми), в том числе, Мукоровые (Mucorales), Эндогоновые (Endogonales), Энтомофторовые (Entomophthorales), Зоопаговые (Zoopagales).
Порядок Гломовые (Glomerales), до недавнего времени входящий в класс Зикомицеты, предложено поднять до уровня типа (отдела) с наименованием тип Glomeromycota (Гломовые).
Характеристика родов
Род Streptomyces
Род Streptomyces – типовой род семейства Streptomycetaceae. Все представители рода – облигатные аэробы. Они нетребовательны к питательным субстратам, не нуждаются в специфических факторах роста, в основном сапрофиты. Стрептомицеты широко распространены в различных типах почв и играют большую роль в процессах минерализации. Важный фитопатоген – Streptomyces scabies, вызывающий обыкновенную паршу картофеля.
Род Streptoverticilliumобыкновенную
Род Streptoverticillium – типовой вид Streptoverticillium baldaccii. Для видов рода характерен ветвящийся субстратный мицелий, диаметром 0,8–1,2 мкм. Воздушный мицелий обычно состоит из длинных прямых гиф с веточками, собранными в мутовки, расположенными через почти равные промежутки. Каждая веточка несет на кончике зонтик из цепочек эллипсоидных или сферических бактериальных спор. Поверхность – от гладкой до немного шероховатой. Цепочки бактериальных спор могут быть прямыми или волнистыми, редко с крючками на концах.
Колонии – мелкие, обособленные, покрыты воздушным мицелием, обычно ватообразной консистенции. Большинство видов синтезируют растворимые пигменты и формируют окрашенный мицелий.
В результате жизнедеятельности представителей рода образуются вещества, обладающие антигрибной, антибактериальной, антипротозойной и антиопухолевой активностью.
Размножаются представители рода фрагментами первичного или вторичного мицелия, либо путем прорастания бактериальных спор.
Представители рода Streptoverticillium – аэробы, хемоорганотрофы, мезофилы, в большинстве почвенные сапрофиты. Субстратный мицелий формируют при низких концентрациях кислорода или при повышении содержания диоксида углерода. Оптимальные условия роста: температура +26°C–+32°C и pH 6,5–8,0.
Род Intrasporangium
Род Intrasporangium – типовой и единственный вид Intrasporangium calvum. Оригинальный штамм выделен при использовании мясо-пептонного агара в чашках Петри, экспонированных на воздухе в школьной столовой. От других аэробных актиномицетов род отличается образованием обильного мицелия и внутримецилярных везикул, составом жирных кислот и фосфолипидов.
Для штаммов рода характерен ветвящийся мицелий, диаметром около 1 мкм, способный распадаться на фрагменты различных размеров и форм. Воздушного мицелия нет. Интеркалярно и на кончиках гиф образуются везикулы лимоновидной или овальной формы, диаметром 5–15 мкм.
Бактерии рода Intrasporangium – грамположительные, некислотоусойчивые, аэробы, хемоорганотрофы, метаболизм окислительного типа, каталозоположительные. Оптимальные условия роста: температура +28°C–+37°C (при +45°C рост прекращается), питательная среда сложного состава, содержащая пептон и дрожжевой экстракт.
Род Kineosporia
Род Kineosporia – типовой вид Kineosporia auranliaca. У колоний, растущих на агаризованной среде, воздушный мицелий отсутствует, имеется центральное возвышение из которого радиально расходятся вегетативные гифы. Иногда на них формируются пучки бактериальных спор в агаре. Зрелые колонии глянцевидные, за счет содержания студенистого матрикса. Бактериальные споры – от сферических до грушевидных или яйцевидных, размером по длинной оси 1–2 мкм. Они образуют цепь вокруг центрального возвышения или располагаются одиночно в виде агрегата на верхушках гиф. За счет полярных жгутиков бактериальные споры данного рода могут передвигаться.
Бактерии рода Kineosporia – грамположительные, аэробы, хемоорганотрофы. Используют простые сахара. Оптимальная температура споробразования и роста +20°C–+30°C. При температуре +37°C роста нет. Местообитание – почва.
Род Sporichthya
Род Sporichthya – типовой вид Sporichthya polymorpha. Воздушный мицелий слаборазветвленный, сформирован короткими редкими гифами, диаметром 0,5–1,0 мкм, растущими с поверхности плотной среды. Воздушные гифы – прямостоячие, поддерживаются выростами стенки базальной клетки. Субстратный мицелий не образуется. Воздушный мицелий разделяется на бактериальные споры от палочковидной до коковидной формы, способные в воде формировать полярные жгутики и становиться подвижными.
Бактерии рода Sporichthya – грамвариабельные. Молодые клетки обычно грамотрицательные, старые в большинстве – грамположительные. По данным электронной микроскопии тип ультратонких срезов клеточной стенки грамположительный.
Биология
Псевдогрибы класса Оомицеты отличаются большим разнообразием в экологическом отношении. В числе видов класса присутствуют сапротрофные и паразитические виды. Последние поражают грибы, водоросли, простейших, ракообразных, насекомых, земноводных, рыб, икру рыб, проростки высших растений.


Обитают представители класса в воде или влажной почве. Чаще всего виды, роды и целые семейства приурочены к определенной среде обитания. Например, виды семейства Lagenidiaceae – внутриклеточные паразиты, Haliphthoraceae – паразиты безпозвоночных, обитающих в пресной и морской воде.
Сапротрофные виды оомицетных грибов развиваются на растительных и животных остатках, но предпочитают водоемы с прозрачной, чистой, стоячей водой. Исключение Leptomituslacteus – обитает в водоемах с отходами различных производств (сахарные заводы, целлюлозные комбинаты). Паразитные формы связаны с организмом-хозяином обычно на уровне вида.
Оптимальная температура развития многих видов +15°C –+18°C. Есть приуроченные к водам с температурой +3°C–+10°C. Большая часть предпочитает умеренные температуры. При более высоких температурных значениях развитие оомицетов подавляется бактериями и другими микроорганизмами. Однако для культивирования в лабораторных условиях оптимальная температура развития оомицетов +20°C–+22°C.
Оптимальное значение pH для многих представителей 7,5–8,0. Отдельные виды выделяются при pH 3,0 (например Saprolegniaturfosa).
Место класса Оомицеты в системе органического мира и макросистеме грибов
Представители класса Оомицеты обладают рядом особенностей морфологического, биологического и химического характера, обосабливающими эту группу организмов от других и заставляющими заявить о неопределенности ее положения в системе органического мира
- Наличие целлюлозы в клеточной стенке, накопление крахмала, присутствие в онтогенезе зооспор, гаметангиальный мейоз – сближает оомицеты с водорослями.
- Строение крист митохондрий, тип организации ферментов синтеза триптофана, молекулярная масса рибосомной РНК, наличие целлюлозы в клеточной стенке – присущи представителям класса Гипохитридиевые (Hyphochytriomycetes).
- Центрическое деление ядер при метозе – сближает оомицеты с хитридиевыми грибами (Хитридиомикотами).
В связи с перечисленными и некоторыми другими особенностями некотые исследователи сближают оомицеты с водорослями, выделяя их в особую группу. В некоторых случаях их рассматривают вместе с хитридиевыми и гипохитридиевыми организмами, либо в царстве Протоктиста (Protoctista) вместе с амебами, водорослями, инфузориями и миксомицетами, или в подцарстве Chromobionta, царства растений.
В настоящее время наиболее достоверным представляется положение типа Оомицеты (Oomycota) в Царстве Хромисты (Chromista) в числе грибоподобных организмов (псевдогрибов), с определением оомикотовые грибы.





