

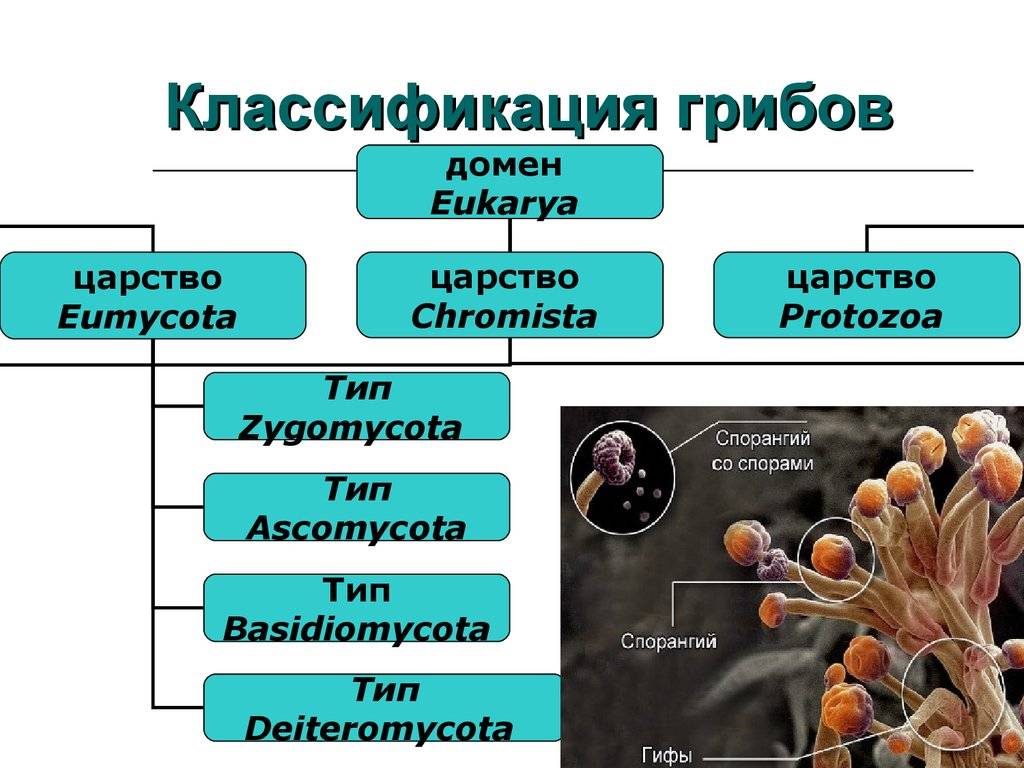
Общие характеристики
Эти грибы являются многоклеточными и образуют неразделенные гифы (псевдоциты). Эти гифы могут расти внутри клеток корня (внутриклеточных) или между ними (межклеточный).
среда обитания
Гломеромикоты распространены по всему миру, занимая практически все биомы планеты. Они имеют тенденцию быть более многочисленными и разнообразными в тропических экосистемах.
Наибольшее количество видов присутствует в Азии, а затем в Южной Америке. Пока в Антарктиде было найдено только три вида.
Они могут присутствовать в нарушенной окружающей среде, связанной с сельскохозяйственными культурами и, в более широком смысле, в наземных природных экосистемах, от тропических лесов до пустынь..
Более 40% видов в этой группе являются космополитическими и только 26% являются эндемичными, в то время как остальные имеют раздельное распределение.
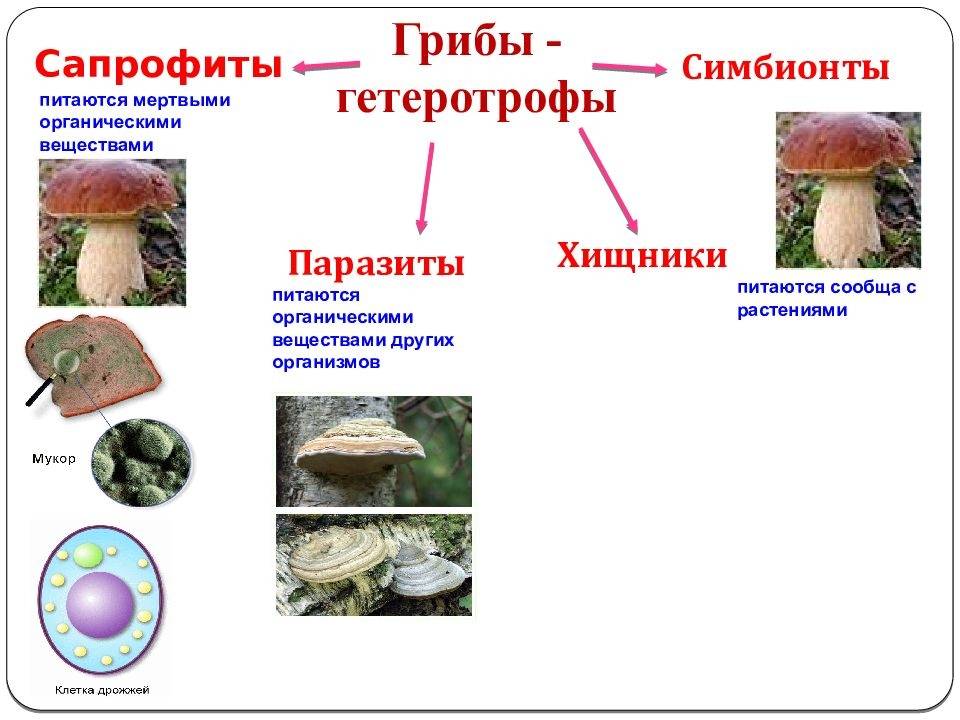
Образ жизни
Гломеромикоты – это облигатные грибы симбионта, то есть они требуют жизни в симбиозе с другими организмами..
Они ассоциируются с корнями растений и образуют эндомикоризы (с грибными гифами в клетках корней растений). Это приносит пользу обоим видам; к грибку и связанному с ним растению.
Грибы, принадлежащие к филломе Glomeromycota, не являются патогенными паразитами, они не вызывают заболеваний или вредных воздействий для других живых существ..
воспроизведение
Грибы Glomeromycota не дают полового размножения. Они размножаются только бесполым путем через хламидоспоры, которые являются спорами устойчивости к неблагоприятным условиям окружающей среды..
Эти грибы рассеиваются в результате фрагментации их мицелия (набор нитей или гиф), а также фрагменты корней растений, которые они колонизировали. Они также рассеиваются через хламидоспоры.
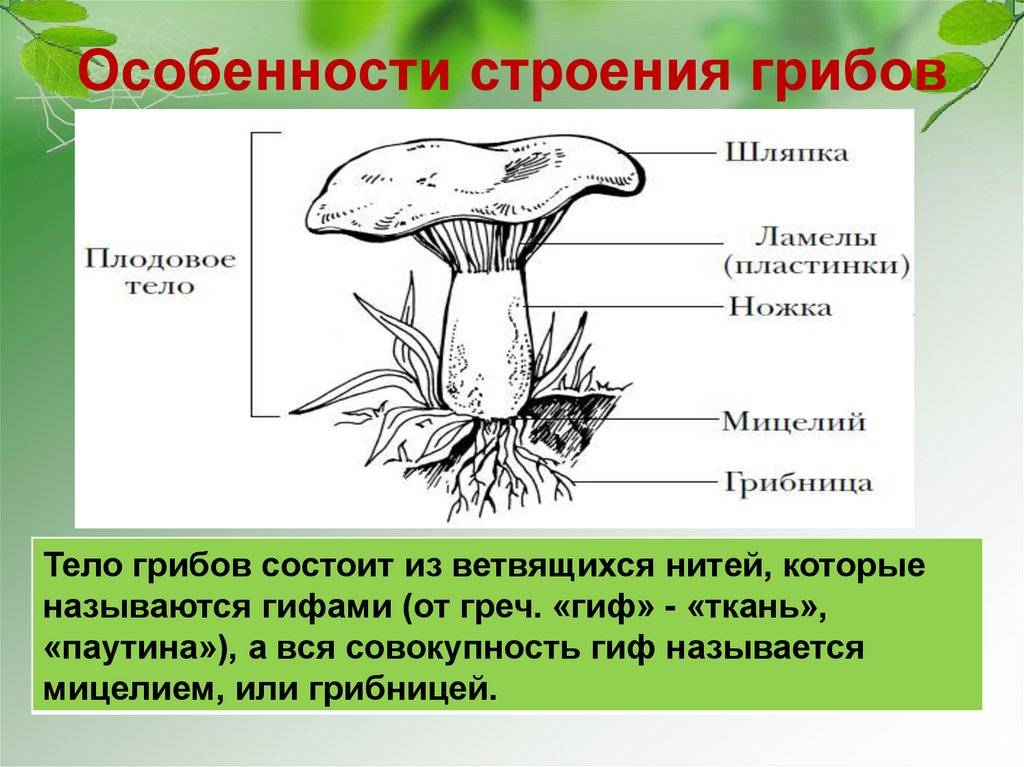
Мицелий и питание
Мицелий или набор нитей грибов Glomeromycota является ценоцитовым; то есть гифы не имеют перегородок или перегородок, а клетки имеют много ядер.
Гифы имеют клеточные стенки с хитином, что придает им жесткость. Такая жесткость и твердость облегчают его проникновение в клетки корней растений..
Мицелий гриба развивается внутри корня (внутрирадикальный мицелий, образующий эндомикоризы), а также вне корня (экстрарадикальный мицелий). Симбиотическая грибно-корневая ассоциация растений называется микоризой.
Грибковые гифы Glomeromycotas также обладают способностью проникать в корковые клетки (или клетки коры, расположенные под эпидермисом) корней и формировать структуры, называемые арбускулы и везикулы..
Арбускулы образованы haustorium или специализированными гифами, которые поглощают питательные вещества из корня растения. Эта хаусториальная гифа очень разветвлена и развивается внутриклеточно (внутри корневых клеток).
Обмен питательными веществами между двумя симбионтами (растение и грибок) происходит в арбускуле.
Гриб снабжает растение макроэлементами, особенно фосфором (P), которые он эффективно извлекает из почвы. Для снабжения растения этими растительными макроэлементами гриб использует экстрарадикальный мицелий, который растет вместе с корнем, но внешне к нему. Растение снабжает гриб сахаром (углеводами), который он вырабатывает благодаря фотосинтезу..
У некоторых грибов Glomeromycota есть пузырьки, которые представляют собой структуры в форме баллона, где они хранят липиды (жиры) в качестве резервных веществ..
Система Hyphas
Система мицелия (набор гиф) состоит из внутреннего мицелия (в тканях корня) и внешнего мицелия (которые распространяются по поверхности почвы)..
Наружные мицелии разветвленные. Они образуют сеть, которая связывает корни растений разных видов в экосистеме.
Во внутренней мицелии есть два типа гиф. Тип Париж они только внутриклеточные и спиралевидные, в то время как аронник они в основном межклеточные.
Внутриклеточные гифы разветвляются, образуя арбускулы (разветвленные гифы, которые занимают более 35% объема инфицированной клетки). Они недолговечны и являются местом обмена питательными веществами между симбионтами..
В некоторых группах Glomeromycota присутствуют пузырьки, которые представляют собой структуры, которые образуются на вершине гиф и накапливают питательные вещества..
Споры бесполые с толстыми и многоядерными стенками. Ядра обычно генетически разные (гетерокариоты).
черты
Все виды рода Moraxella аэробные, неподвижные, имеют фимбрии, не продуцируют пигмент или гемолиз на агаре крови.
Они положительные оксидаза и каталаза, важные тесты для дифференциации рода Moraxella других родов морфологически очень похожи. Например, оксидаза помогает отказаться от рода Acinetobacter, и каталаза исключает род Kingella.




Другой род, с которым их можно спутать, особенно вид М. catarrhalis, Это с Neisseria, как по морфологии, так и по оксидазному тесту.
В этом случае они различаются по неспособности пола Moraxella образовывать кислоту из углеводов, в то время как большая часть Neisseria если они способны ферментировать некоторые углеводы.
Еще один способ дифференцировать пол Neisseria рода Moraxella выполнить окраску по Граму, взяв колонии от внешнего края гало ингибирования вокруг диска пенициллина.
Виды Neisseria полностью кокосовое и те из Moraxella они будут производить удлиненные, плеоморфные формы. Со своей стороны, М. фенилпирувица характеризуется гидролизом мочевины и дезаминированием фенилаланина.
Moraxella catarrhalis характеризуется продуцированием ДНКазы, гидролизом трибутирина и восстановлением нитратов до нитритов.
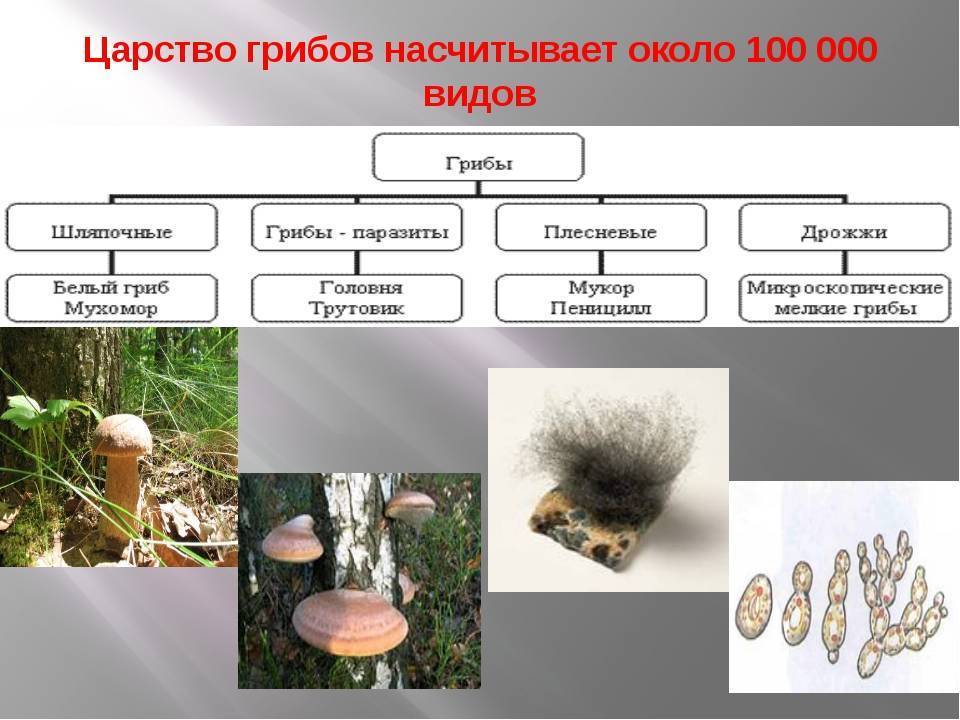
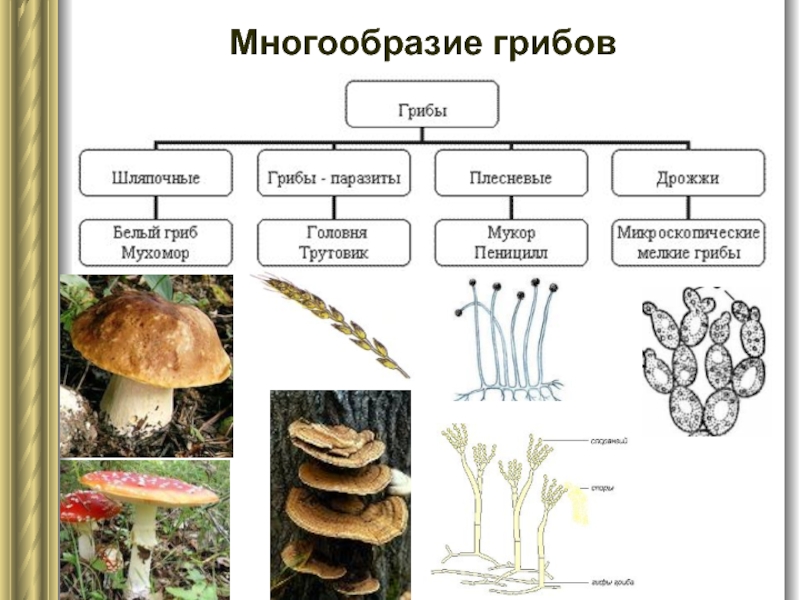
Примеры грибов Glomeromycota: род гломусной
Среди грибов Glomeromycota можно выделить несколько видов, принадлежащих к роду Glomus, которые являются родом микоризных арбускулярных грибов (AM), с видами, которые образуют симбиотические ассоциации (так называемые микоризы) с корнями растений. Это самый многочисленный род грибов AM из 85 описанных видов..
Среди видов рода Glomus можно выделить: Glomus aggregatum, G. mosseae. G. flavisporum, G. epigaeum, G. albidum, G. ambisporum, G. brazillanum, G. caledonium, G. coremioides, G. claroideum, G. clarum, G. clavisporum, G. constrictum, G. coronatum, G. дезертикола, G. diaphanum, G. eburneum, G. etunicatum, G. macrocarpus, G. intraradices, G. microcarpus, Г. незначительный, среди других.
ссылки
- Koneman E, Allen S, Janda W, Schreckenberger P, Winn W. (2004). Микробиологический диагноз. (5-е изд.). Аргентина, редакция Panamericana S.A..
- Forbes B, Sahm D, Weissfeld A. Bailey & Scott Microbiological Diagnosis. 12 изд. Аргентина. Редакция Panamericana S.A; 2009.
- Гонсалес М., Гонсалес Н. Руководство по медицинской микробиологии. 2-е издание, Венесуэла: Управление средств массовой информации и публикаций Университета Карабобо; 2011
- Гомес-Камараса C, Фернандес-Парра J, Наварро-Мари J, Гутьеррес-Фернандес Й. Возникающая инфекция Moraxella osloensis. О генитальной инфекции. Преподобный Esp Quimioter, 2018; 31 (2): 178-181
- Отазо Д., Инохоса М., Сильвия А., Хомси Мальдонадо, Надя Ю., Поцци Г. Антибиограмма и распространенность Moraxella catarrhalis в лаборатории “Институт патологии Кочабамба 2005-2010. Преподобный Сиен Мед. 2014; 17 (1): 23-25.
- Esparcia O, Magraner J. Moraxella catarrhalis и его значение в инфекционной патологии. Служба микробиологии. Университетская клиническая больница Валенсии. pp1-9
- Участники Википедии. Moraxella. Википедия, Свободная энциклопедия. 22 марта 2018 года, 13:42 UTC. Доступно по адресу: en.wikipedia.org
- Ян М, Джонсон А., Мерфи Т.Ф. Характеристика и оценка Moraxella catarrhalis олигопептид-пермеаза А как антиген слизистой вакцины. Заражение Иммун 2010; 79 (2): 846-57.
Морфология
Имаго. Комар размером 6–8 мм. Морфология вида подвержена значительным колебаниям по окраске и величине. Экземпляры, обитающие на юге, характеризуются более светлой окраской и меньшими размерами.
Голова с боков усажена торчащими темно-бурыми чешуйками, сверху расположен направленный вперед пучок длинных беловатых чешуек и волосков. Усики бурые. Щупики и хоботок бурые, преимущественно в прилегающих чешуйках.
Среднеспинка в средней части с широкой, кпереди суживающейся продольной полосой сероватого цвета. На передней половине данной полосы расположены, как правило, 2–3 неясные буроватые полоски. По переднему краю среднеспинки у головы располагается пучок беловатых чешуек. Боковые отделы в передней части бурые, в задней – черно-бурые. Волосяной покров в средней части и на щитке светло-бурый, в остальной части, особенно над основаниями крыльев, темно-бурый.
Ноги бурые, с нижней стороны бедер и внутренней стороны голеней более светлые, лапки темные.
Крылья покрыты чешуйками буроватого оттенка, с черными пятнами более темных черно-бурых чешуек, расположенных при основании радиальной и медиальной вилок. Чешуйки по краю крыла и на его вершине светло-желтые.
Жужжальца желтые, с буроватой головкой.
Брюшко бурое или черно-бурое, часто с более светлыми пятнами на боковых отделах тергитов, покрыто довольно длинными золотисто-желтыми или буроватыми волосками.
Половой диморфизм. Разнополые особи отличаются строением половых органов.
Самка. Хоботок состоит из верней и нижней губы, подглоточника, пары верхних и пары нижних челюстей. Щупальца длинные, примерно равны хоботку.
Самец. Парные части хоботка редуцированы. Щупальца также же длинные, но с явственно утолщенными конечными члениками.
Яйцо оборудовано плавательной камерой, занимающей среднюю треть яйца, и оторочкой. Структура и окраска экзохориона различается, но характерна для отдельных подвидов. Она зависит от местных изменений строения наружной оболочки и состоит из светлых и темных пятен и полос.
Личинка IV стадии. По причине большого биологического разнообразия мест обитания и условий развития личинки вида изменчивы по внешнему виду, величине и окраске. Особи северных широт отличаются более крупными размерами и более темной пигментацией на хитиновых образованиях (голове, стигмальных пластинках и пр.).
Общая окраска тела варьирует от серо-желто-зеленой до черно-зеленой.
Пигментация головы изменчива – от светло-коричневых пятен до черно-коричневых полос на общем темном фоне.
Усики почти прямые, до 2/3 длины головы, с более темным концом. Волосок короткий, в среднем равен ширине усика у места своего отхождения, лежит у основания и состоит из 4–6 тонких веточек.
Стигмальная пластинка крупная. Твердые хитиновые образования сильно пигментированы. Передняя лопасть с темным основанием, остальная часть заметно светлее. Стигмы на некотором расстоянии от центральной пластинки отростков не имеют.
Гребень на боковых пластинках VIII членика состоит из 6–10 (чаще 7–9) крупных шиповидных зубцов, между ними находятся от 1 до 4 более мелких зубцов.
Плавник составлен из 18–20 пучков. Жабры в среднем равны длине седла, обе пары одной длины.
Куколка, как и у всех представителей рода Anopheles, сбоку имеет вид запятой, сверху – неправильного овала. Тело состоит из двух отделов: массивной головогруди и тонкого уплощенного дорсовентрального брюшка. Глаза сильно пигментированы и просвечивают через прозрачную кутикулу. Над ними по бокам спинки отходят куколочные рожки, суженные у основания и сильно расширяющиеся к свободному концу. Их отверстия скошены внутрь и при нахождении куколок на поверхности лежат в плоскости поверхностной пленки. От основания рожек отходит трахея, соединяющая полость рожек с переднегрудной стигмой развивающегося комара.
Брюшко куколки состоит из девяти члеников. На заднем крае спинной стороны VIII членика находятся два плавника, заходящие друг за друга внутренними краями. Каждый плавник имеет вид неправильной овальной пластинки, сидящей на суженном основании. Пластинка укреплена продольным срединным ребром, не доходящим до ее свободного края. От конца ребра отходит волосок, у его основания располагается более короткий добавочный волосок.




морфология
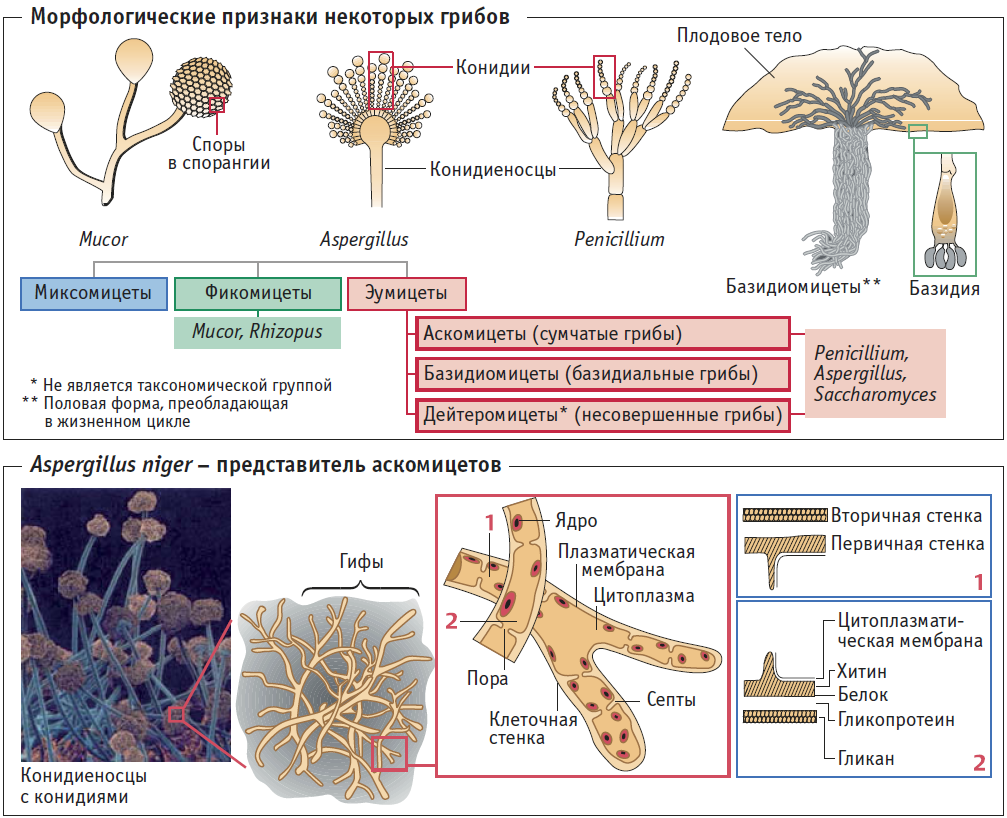
Пол Moraxella в граме окраска может рассматриваться как диплобациллы, коккобациллы или грамотрицательные диплококки, в зависимости от вида.
В частном случае Moraxella catarrhalis, единственный вид, имеющий грамположительную морфологию диплококков.
Макроскопически после 24 часов инкубации на кровяном агаре наблюдаются небольшие и точечные колонии диаметром менее 0,5 мм, серого цвета..
С другой стороны, большинство штаммов рода Moraxella растут с трудом и медленно на агаре Макконки с неферментирующими лактозными (бледными) колониями, тогда как другие просто не растут как М. лакуната и М. nonliquefaciens.
Некоторые виды
Вот некоторые виды рода Moraxella и его макроскопические или культурные характеристики.
Moraxella atlantae он медленно развивается в питательных средах и производит колонии с тенденцией к образованию инвазивной зоны после 48 часов инкубации.
Со своей стороны, Моракселла Лакуната Требуются особые требования к выращиванию в виде обогащенной среды без пептона, добавления олеиновой кислоты или кроличьей сыворотки для противодействия токсическому протеолитическому эффекту. Растет на кровяном агаре в виде укуса.
Moraxella nonliquefaciens также необходимы сывороточные добавки для оптимального развития и выделяются, потому что их колонии очень слизистые.
Moraxella osloensis и M. phenylpyruvica им не нужны особые требования к их изоляции.
Моракселла Канис образует большие, гладкие колонии на агаре с кровью баранов, аналогичные колонии семейства Enterobacteriaceae.
воспроизведение
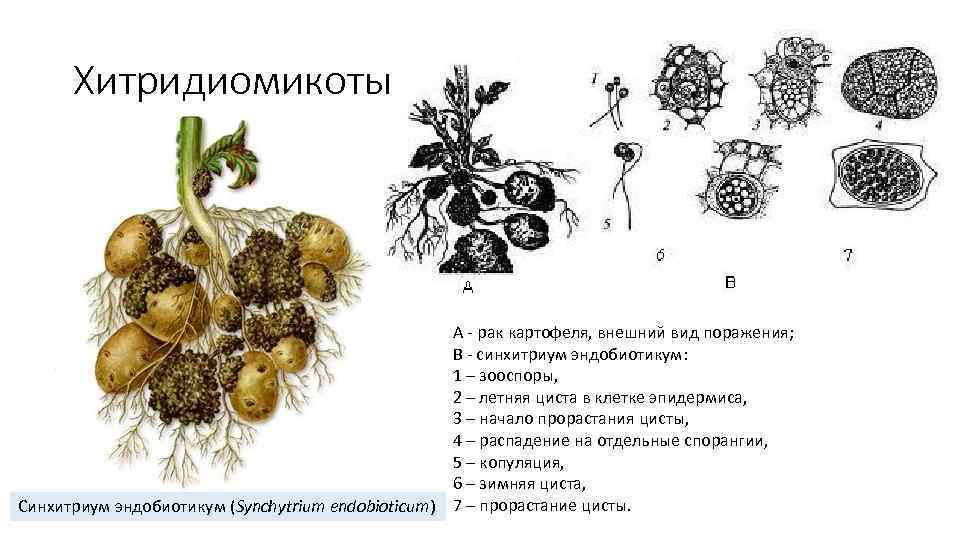
До настоящего времени в Glomeromycota было зарегистрировано только бесполое размножение..
Бесполые споры имеют очень толстую стенку и большие (40-800 мкм). Это может произойти в sporocarp (сеть гиф), которая формируется непосредственно в корне, почве или других структурах (остатки семян, насекомых или других). Они многоядерные (от сотен до тысяч ядер) и могут быть генетически отличимыми
Колонизация хозяина
Споры падают на землю и переносятся насекомыми, мелкими млекопитающими или водой. Позже они прорастают, проходя через очень короткую фазу сапрофита. Проросшие трубки могут расти на 20-30 мм до колонизации корня..
Как только герминативная трубка вступает в контакт с корнем, образуется апрессор (адгезивная структура), который проникает в клетки эпидермиса. Гифы достигают радикальной коры, как внутриклеточно, так и внутриклеточно, и образуются arbúsculos, пузырьки и внерадикальная сеть гиф.
Морфологически близкие виды
По внешнему виду (морфологии) имаго близок Комар малярийный камышовый (Anopheles hyrcanus). Отличается наличием на переднем крае крыльев светлых чешуек, образующих два пятна, и торчащими чешуйками щупиков.
Кроме того, отмечается, что вид Anopheles maculipennis разделяется на несколько рас, иногда они трактуются как подвиды. Шесть из них представлены в фауне Палеарктики: maculipennis, messefe, melanoon (subalpinus), labranchiae, atrorparvus, sacharovi.
Высказываются предложения о возведении названных рас в ранг видов. Однако такая крайняя точка зрения общего признания не получила, поскольку формы комплекса maculipennisближе друг к другу, чем к другим формам. Намечающиеся группы внутри комплекса иногда предлагается рассматривать в качестве видов. Такая трактовка принята многими, в частности авторами всесветного каталога комаров. В составе фауны Палеарктики вместо одного вида Anopheles maculipennis признаются три: maculipennis с подвидами maculipennis, messefe, melanoon; labranchiae с подвидами labranchiae и atrorparvus; и sacharovi.
Североамериканский Аn. occidentalis тоже рассматривается в качестве отдельного вида. Данные четыре вида совместно с некоторыми другими близкими и образуют группу maculipennis, которая входит в состав одной из шести групп видов, на которые подразделяется подрод Anopheles.
Сторонники дробления комплекса maculipennis на большее или меньшее количество видов указывают на различие в дорсальной поверхности яиц и наличие биологических отличий между формами.
Однако не обнаружено никаких достоверных различий между членами комплекса ни по личинкам, ни по имаго, ни по строению гипопигия (основным принципам классификации комаров). Известны только некоторые отличия sacharovi, но они несущественны и имеют характер отличий только подвидового ранга.
Что касается биологических особенностей, отмечается, что довольно существенные внутривидовые различия выявляются у многих других видов комаров. В этом случае имеют дело, скорее всего, с биотипами. Поэтому вопрос о дроблении как maculipennis, так и других видов, признать оправданным нельзя.
Мнение о существовании строгой репродуктивной изоляции между членами комплекса также не находит твердого опытного подтверждения.
Филогения и таксономия
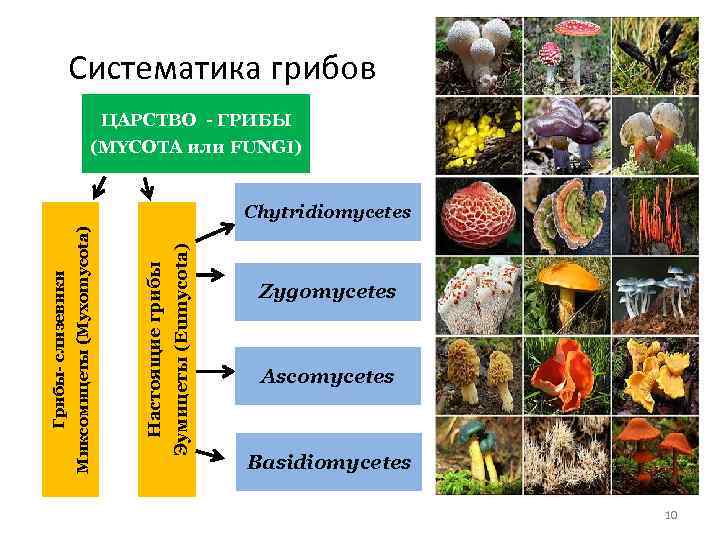
Первые Glomeromycota были обнаружены в 19 веке и были обнаружены в классе Zygomycetes из-за наличия толстостенных спор. В течение 1990-х годов было установлено, что все грибы арбускулярной микоризы были облигатными симбионтами с уникальными морфологическими характеристиками..
В 2001 году край Glomeromycota создан на основе морфологических, биохимических и молекулярных характеристик. Это братская группа под царства Дикарья.
заказы
Он разделен на четыре категории: археоспоралы, диверсиспоралы, гломералы и парагломералы. Они включают 13 семейств, 19 родов и 222 вида..
Археоспоралы образуют эндосимбионты с цианобактериями или микоризы с арбускулой, а их споры бесцветны. Он состоит из трех семейств и примерно пяти видов.
Diversisporales представляют арбускулы и почти никогда не образуют пузырьки. Восемь семей и около 104 видов были описаны.



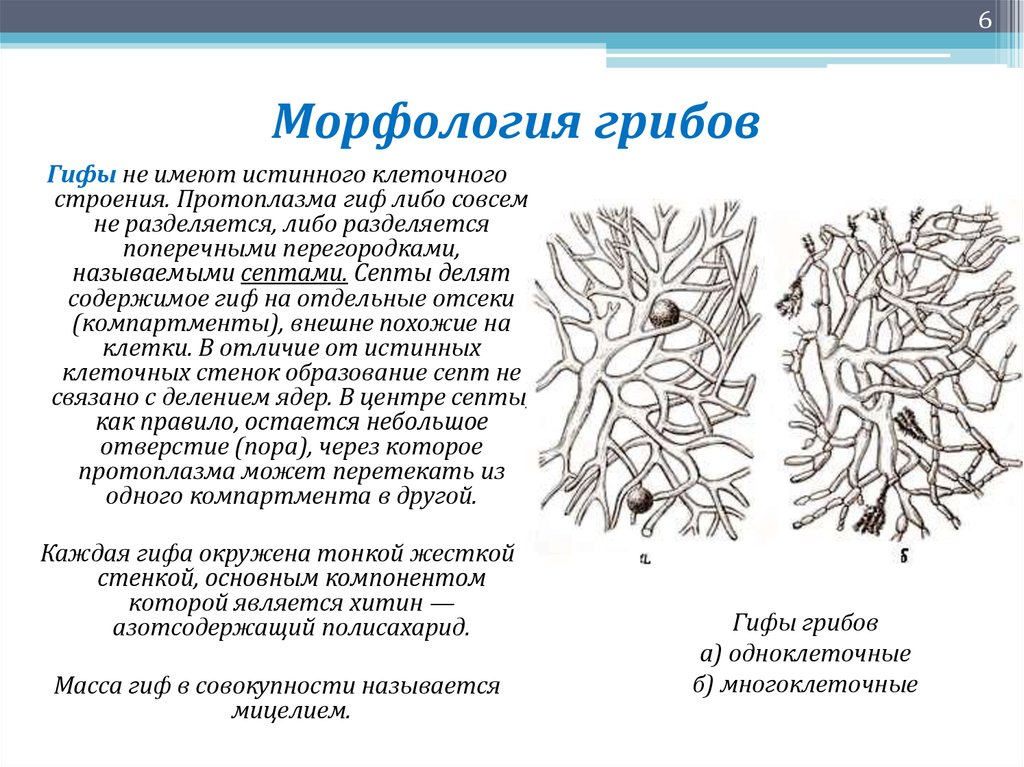
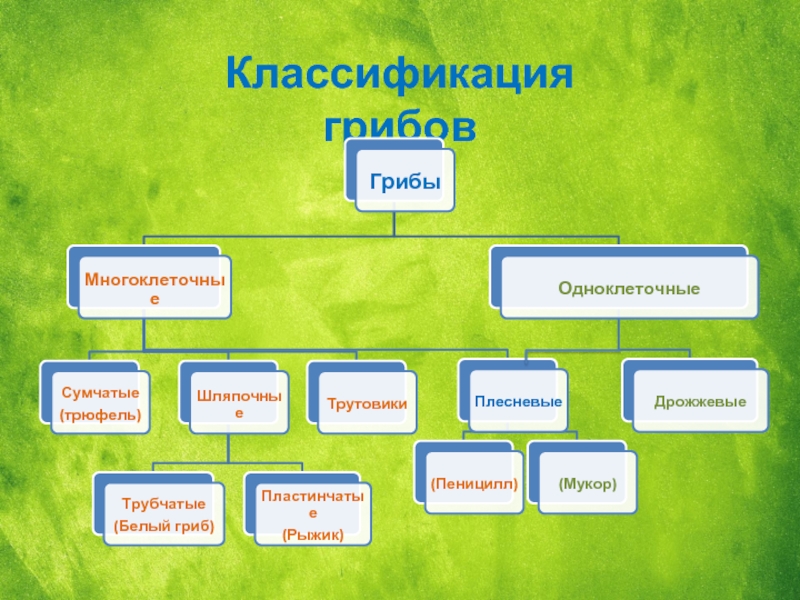
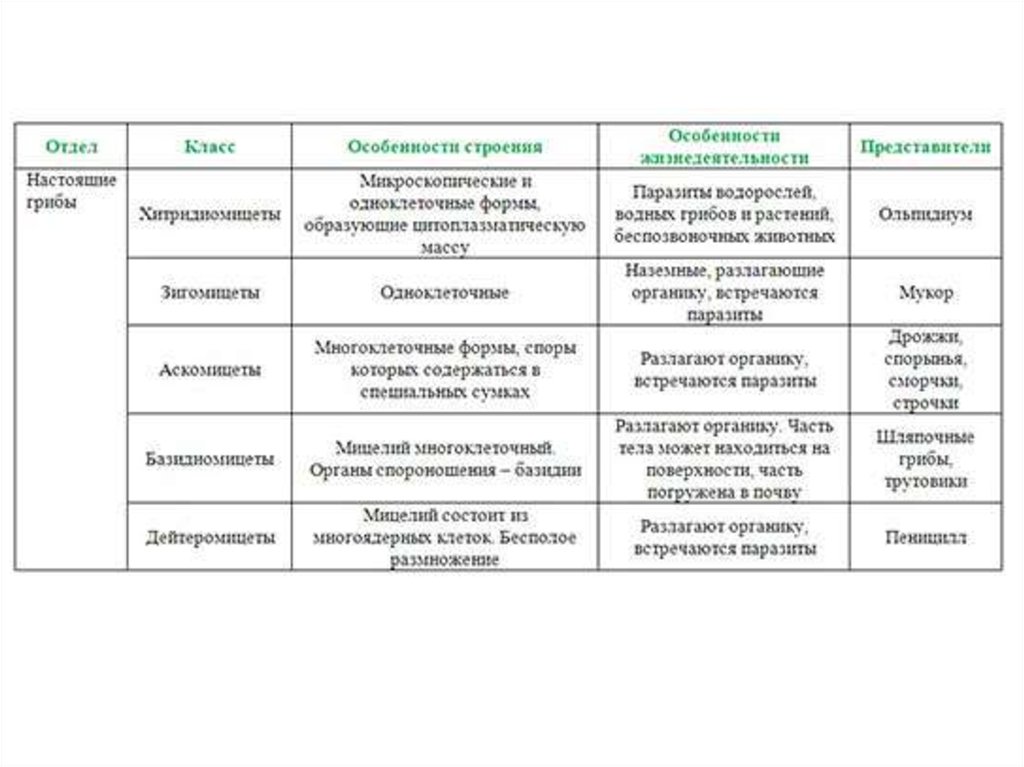
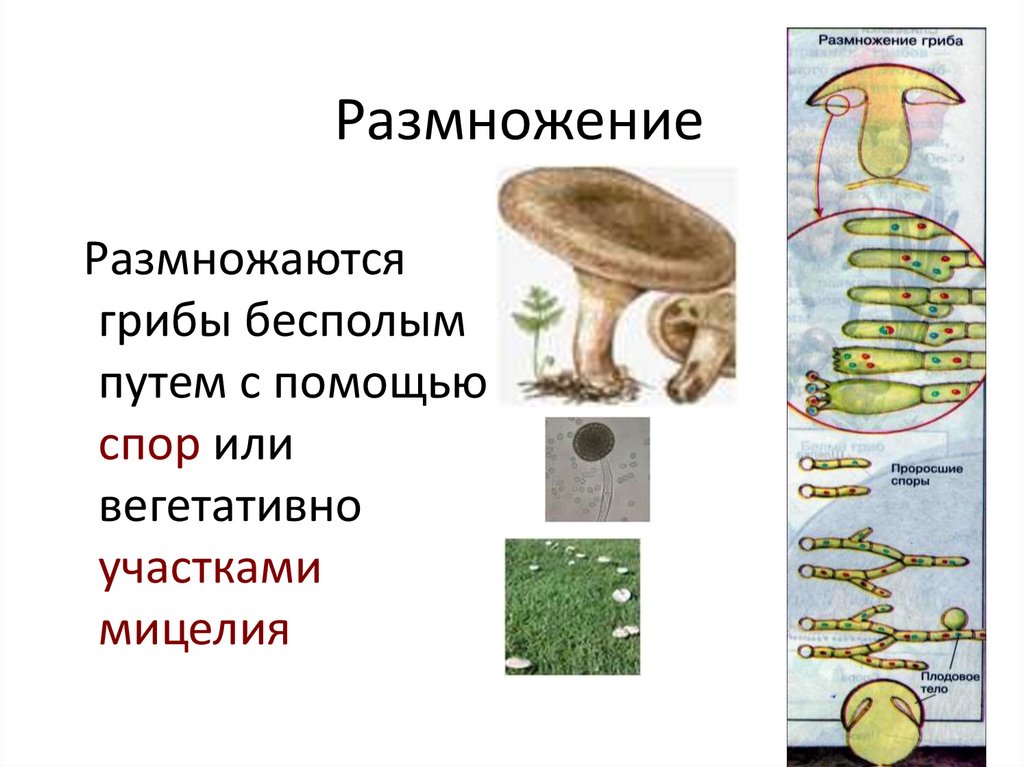
Glomerales – самая большая группа. Он представляет арбускулы, везикулы и споры с различной морфологией. Он состоит из двух семей и пола гломусной Это самый большой около 74 видов.
В парагломералах арбускулы присутствуют, пузырьки не развиваются, споры бесцветны. Это содержит семью и род с четырьмя описанными разновидностями.
Таксономия
Marasniellus Candidus (Bolton) Певец
Marasmiellus представлял только три вида, когда он был впервые описан Уильямом Мурриллом ( Marasmiellus inconspicous Murrill (Куба), Marasmiellus purpureus (Berk. & Curt.) Murrill (Куба) и Marasmiellus juniperinus Murrill (Ямайка)). В настоящее время роды насчитывают более 250 видов, из которых типовой вид M. juniperinus остается таким же, как и при первом описании родов. Marasmiellus традиционно был включен в Tricholomataceae R. Heim ex Pouzar, но более поздние исследования ДНК показали, что части этого рода принадлежат к Omphalotaceae , который был описан А. Бресинским в 1985 году и считается синонимом Marasmiaceae . Более того, несколько исследований показывают, что Marasmiellus разветвляются на несколько полифилетических и искусственных групп . Это привело к конфликту вокруг его положения относительно Gymnopus и привело к двум противоположным взглядам на их таксономию: 1) Marasmiellus входит в состав Gymnopus (включительно) или 2) Marasmiellus остается отдельным от Gymnopus родом (ограничительный). Самые последние исследования показали, что вторая гипотеза является наиболее сильной, выделяя Marasmiellus в отдельный род, но для подтверждения этого требуются дополнительные исследования.
Развитие
Имаго. Зимует во взрослой фазе оплодотворенной самки как эндофильный вид в различных нежилых помещениях, часто совместно с сельскохозяйственными животными. В природе – в пещерах, норах. В северной и восточной частях ареала комары выносят многомесячное воздействие очень низких температур. Температурный минимум развития составляет +7–10 °С.За теплое время года вид дает 2 поколения на севере и 4–6 на юге. Максимум численности наблюдается в середине лета, при очень жаркой погоде отмечается снижение численности.
Главная масса комаров концентрируется в населенных пунктах или недалеко от них. Питаются в основном кровью сельскохозяйственных животных, домашних и диких птиц, в меньшей степени людей, но при отсутствии животных основной источник кровососания – люди. Питание кровью необходимо самкам. Автогенного развития яиц у данного вида не наблюдается. Для данного вида характерна гонотрофическая гармония – одного полного насыщения кровью достаточно для развития и откладки яиц.
Период спаривания. Копуляция проходит на лету при роении. Яйца в теле самок при температуре +7,5°С созревают в течение 23 суток, при +1 °С – 7 суток, а при +20°С – 4 суток. При оптимальной температуре +25–30°С – в течение 2–3 суток. После созревания яиц самки вылетают с мест дневок и ищут водоемы для откладывания яиц. Самки откладывают яйца поодиночке, впоследствии те могут сцепляться друг с другом, но не склеиваться. Количество яиц колеблется от 100 до 500 штук.
Яйцо. Как у всех кровососущих комаров, длительность эмбрионального развития может колебаться от 40 часов до 8 суток, при температуре +23–24°С – 3–6 суток.
Личинка. Места обитания – разнообразные по типу, глубине и величине водоемы, хорошо освещенные солнцем на севере и затененные на юге. В южных районах обитают в слабо солоноватой воде. Оптимальная скорость развития наблюдается при температуре +29–30°С – 5–7 суток, при +15–16°С – до 30 суток.
Куколка. Зрелая личинка IVстадии после линьки переходит в куколку. Куколки не питаются и с внешней средой связаны только процессами газообмена. В нормальном состоянии они свешиваются вниз с поверхностной пленки воды. Куколки способны к быстрым активным движениям и опускаются на дно водоема при внешних раздражениях. В спокойном состоянии куколки долго находятся на поверхности воды. Длительность куколочной фазы в среднем не превышает 2–2,5 суток. Первыми всегда вылетают самцы.
Морфология и жизненный цикл
Морфология Marasmiellus уделяется мало внимания по сравнению с другими родами Omphalotaceae , в основном из – за их uncolorful шляпки , маленькие плодовые тела , и мало вариации морфологических признаков. Эти факторы затрудняют разграничение видов внутри этого рода. Виды Marasmiellus имеют распростертые и дивертикулированные гифы , которые не имеют четкой ориентации. Однако было замечено, что внешние гифы могут собираться в пучки и быть радиально ориентированными. Кроме того, хейлоцистидии возникают из горизонтальных гиф и часто внедряются в гимений , часто оказываясь ниспровергнутыми. Некоторые виды Marasmiellus используют прорастание базидиоспор , и различение разных родов на основе воспроизводства будет зависеть от скорости прорастания. Во время прорастания споры Marasmiellus имеют тенденцию рассеиваться на чашках для разведения. Некоторые виды тетраполярны. Кроме того, ни о каких биполярных или амфиталлических таксонах в этом роде не сообщалось, что является неожиданным, поскольку такое поведение при спаривании является обычным для тропических агариков , позволяя быстро колонизировать. Их базидиокарпы бывают коллибиоидными или омфалиоидными и имеют белый отпечаток спор. Кутис consis из pileipellis , который иногда переходит в trichoderm, с или без Rameales-структуры.
Чувствительность к антибиотикам
Поскольку эти виды были очень чувствительны к пенициллину, антибиограмма не требовалась. Тем не менее, известно, что с 1990 года большинство штаммов, особенно видов catarrhalis, являются производителями бета-лактамаз.
Поэтому желательно провести тесты на чувствительность к противомикробным препаратам. Но эту работу сложно выполнить, поскольку в CLSI не указаны точки среза для жанра. Moraxella, что мешает его интерпретации.
По этой причине в некоторых лабораториях используются точки разреза, описанные для Haemophilus influenzae или для беспокоящих бактерий, которые могут расти в средах Мюллера-Хинтона без добавок, или для неферментирующих грамотрицательных бактерий.
Штаммы Moraxella catarrhalis часто чувствительны к хинолонам, амоксициллин-клавулановой кислоте, цефалоспоринам, тикарциллину, пиперациллину, макролидам, хлорамфениколу и аминогликозидам.
Однако сообщалось о штаммах, устойчивых к тетрациклину, эритромицину, фторхинолону, макролидам, пиперациллину и некоторым цефалоспоринам..





